线粒体DNA
粒線體DNA(英語:,縮寫作)指一些位於粒線體內的DNA,與一般位於細胞核內的DNA有不同的演化起源,可能是源自早期細菌。雖然現存生物體中絕大多數作用於粒線體的蛋白質,是由細胞核DNA所製造,但這些基因中有一些可能是源於細菌,並於演化過程中轉移到細胞核中,稱為核內線粒體片段。
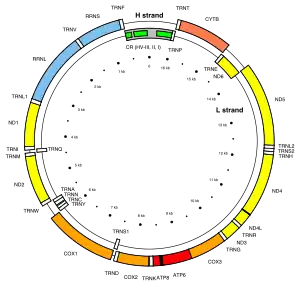
現今人類體內的每個細胞中,大約有1000到10000個粒線體,而每一個粒線體內,則大約有2到10組mtDNA,每個mtDNA共包含16569個鹼基對,其中有37個基因,可用來製造13種蛋白質、22種tRNA與兩種rRNA。其中的內含子較細胞核基因少,且有些不含內含子,如tRNA基因。
突變速率
動物體內的粒線體DNA並不會經過遺傳重組,因此與細胞核DNA相較之下有較高的突變速率(重組有修復突變的功能);而植物與真菌類體內的mtDNA則存在著重組現象,其中植物的mtDNA突變速率高於細胞核DNA;真菌方面的mtDNA突變速率則尚未明瞭[2]。
由於動物的mtDNA的突變速率高於細胞核DNA,較為容易測量計算,使mtDNA成為用來追溯動物母系族譜的有效工具,例如許多物種在數百個世代以前的祖先。此外,人類的mtDNA也可用來進行個體辨識。
粒線體DNA起源
內共生理論認為真核細胞最早的起源,是因為原核細胞(細菌等)被吸收到另外一個細胞中,而沒有被消化。而這兩個細胞之後產生了共生關係,使最早的細胞器誕生,此胞器後來成為現今的粒線體,其基因組也在演化過程中轉變成粒線體DNA。
粒線體遺傳
對動物而言,受精卵中的mtDNA主要遺傳自母親;而對植物來說略有變異,但仍然以母系遺傳為主;真菌則源自雙親。
雌性遺傳
正常狀況下,粒線體只會遺傳自母親,以哺乳類而言,一般在受精之後,卵子細胞就會將精子中的粒線體摧毀[3]。1999年發表的研究中顯示,父系精子粒線體(含有mtDNA)帶有泛素(ubiquitin)標記,因而在胚胎中會被挑選出來,進而遭到摧毀[4]。不過某些細胞外的人工受精技術可直接將精子注入卵子細胞內,可能會干擾摧毀精子粒線體的過程。
由於母系遺傳的特性,使得研究者能夠藉由粒線體DNA,追溯長時間的母系族譜(與之相對的為專門用來追溯父系族譜的Y染色體)。人類的粒線體DNA中累積了一些高度變異控制區域(hypervariable control region;包括HVR1與HVR2),在HVR1中含有大約440個鹼基對,這些鹼基對可用來與其他個體(特定人士或資料庫中的訊息)的控制區作比對,進而測定出母系族譜。Vilà等人的研究中回溯了家犬與狼的母系祖先,同樣的分析方式也導出了粒線體夏娃概念,用於研究人類起源。
由於mtDNA並非高度保守,而是擁有較快的突變速率,因此可用來研究種系發生學,生物學家挑選少量不同物種的基因,分析其序列的保留與變異程度,可建立出演化樹。
遺傳疾病
粒線體DNA的突變可造成許多的疾病,如運動障礙(exercise intolerance)或凱塞症候群(Kearns-Sayre syndrome,KSS),一種可使患者心臟、眼睛與肌肉完全失去运动功能的疾病。
遺傳密碼
真菌、原生生物和人以及其它动物的粒線體中的遺傳密碼與標準遺傳密碼的差異,主要變化如下:
| 密碼子 | 通常的作用 | 例外的作用 | 所属的生物 |
| UGA | 中止編碼 | 色氨酸編碼 | 人、牛、酵母线粒体,支原体(Mycoplasma)核基因组,如Capricolum |
| AGR | 精氨酸編碼 | 中止編碼 | 大部分动物线粒体,脊椎动物线粒体 |
| AGA | 精氨酸編碼 | 丝氨酸編碼 | 果蝇线粒体 |
| AUA | 異亮氨酸編碼 | 蛋氨酸編碼 | 一些动物和酵母线粒体 |
| AAA | 赖氨酸編碼 | 天冬氨酸編碼 | 一些动物的线粒体,果蝇线粒体 |
| CUN | 亮氨酸編碼 | 苏氨酸編碼 | 酵母线粒体 |
參考文獻
- Iborra FJ, Kimura H, Cook PR; Kimura; Cook. . BMC Biol. 2004, 2: 9. PMC 425603. PMID 15157274. doi:10.1186/1741-7007-2-9.
- Scott Freeman & Jon C. Herron. Evolutionary Analysis 3rd edition. Pearson Education. ISBN 0-13-101859-0
- 為什麼線粒體DNA不能通過父系遺傳?
- Sutovsky, P., et. al. . Nature. Nov 25, 1999, 402: 371–372. doi:10.1038/46466. Discussed in .
- Mitochondria can be inherited from both parents
- Kondo R, Matsuura ET, Chigusa SI (1992). Further observation of paternal transmission of Drosophila mitochondrial DNA by PCR selective amplification method. Genet Res 59: 81-84.
- Meusel MS, Moritz RF (1993). Transfer of paternal mitochondrial DNA during fertilization of honeybee (Apis mellifera L.) eggs. Curr Genet 24: 539-543.
- Gyllensten U, Wharton D, Josefsson A (1991). Paternal inheritance of mitochondrial DNA in mice. Nature 352: 255-257.
- Shitara H, Hayashi JI, Takahama S, Kaneda H, Yonekawa H (1998). Maternal inheritance of mouse mtDNA in interspecific hybrids: segregation of the leaked paternal mtDNA followed by the prevention of subsequent paternal leakage. Genetics 148: 851-857.
- Zhao X, et al. (2004). Further evidence for paternal inheritance of mitochondrial DNA in the sheep (Ovis aries). Heredity 93:399-403.
- Steinborn R, Zakhartchenko V, Jelyazkov J, Klein D, Wolf E, Muller M et al (1998). Composition of parental mitochondrial DNA in cloned bovine embryos. FEBS Lett 426: 352-356.
- Schwartz M, Vissing J (2002). Paternal inheritance of mitochondrial DNA. N Engl J Med 22: 576-580.
- Daniel Hartl & Elizabath W. Jones. Genetic. 6th ed. Jones and Bartlett Publishers. ISBN 0-7637-1511-5