原杉藻
原杉藻(學名:)為一屬陸生的真菌,生存於志留紀晚期至泥盆紀晚期(4億3千萬年前 - 3億6千萬年前),其子實體形成類似樹幹的結構,直徑達1(3英尺),高度則可達8.8(29英尺)[1],由直徑50微(0.00016英尺)的管束交織而成,為當時陸地上最大的陸生生物。
| 原杉藻属 化石时期:晚志留紀 - 晚泥盆紀 | |
|---|---|
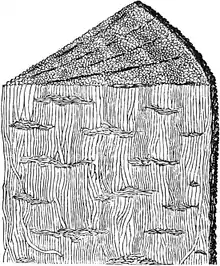 | |
| 原杉藻枝幹的描繪(1888年) | |
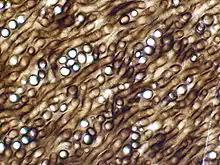 | |
| 光學顯微鏡下的原杉藻組織切片 | |
| 科学分类 | |
| 界: | 真菌界 Fungi |
| 科: | †原杉藻科 Prototaxitaceae |
| 属: | †原杉藻属 Prototaxites Dawson, 1859 |
| 種 | |
| |
| 異名 | |
| |
傳統上很難將原杉藻歸類至現存的分類系統中,目前的觀點認為它們屬於真菌[1],也有研究認為它們為藻類的共生體(即地衣),且其組織中的管束細胞與球囊菌門的十分相似[2]。而對於原杉藻與其他真菌類群的關係亦有多種觀點,除了認為其與球囊菌門接近者外,也有觀點認為其屬於擔子菌門、毛黴門或子囊菌門[3]。
形態學

原杉藻化石主幹的直徑達1(3英尺),高度可達8.8(29英尺)[1],為當時陸地上最大的陸生生物。它們主幹的基部較粗且具有分枝,可能具有類似根部的構造,但仍然未有化石紀錄[4]。可能為原杉藻根部構造的鑄型印痕化石在泥盆紀地層年代十分常見[5]。主枝幹具有環狀生長輪,這些生長輪中偶爾會嵌入一些植物殘骸[6],顯示原杉藻可能透過不斷包覆外層來進行成長。它們的主枝幹可能為子實體,透過菌絲體供給營養。微觀上,原杉藻的枝幹由兩種管束(可能為菌絲)交織而成:骨架管束直徑約20至50微(6.6×10−5至0.000164英尺),內壁厚度約2至6微(7.9×10−5英寸至0.000236英寸);另一種增生管束則較細,直徑約5至10微(1.6×10−5至3.3×10−5英尺),且具有許多交叉的分枝,增生管束內具有隔膜,且隔膜上具有小孔隙,這項特徵只有在現存的真菌與紅藻上發現[7]。
志留紀晚期疑似為植物的化石織絲體同樣有發現類似的管束結構,顯示織絲體可能其實為原杉藻上的葉狀組織,然而這項理論仍然處在假說階段,缺乏相關支持證據[8]。
研究歷史
原杉藻的化石最早發現於1843年[9],然而一直到了14年後才由加拿大地質學家約翰·威廉·道森開始研究原杉藻的化石並進行發表,最初道森認為這個樣本屬於已經部分腐爛的松柏門植物組織,並且上面附生有正在分解組織的真菌[6]。1872年,蘇格蘭植物學家威廉·卡魯瑟斯質疑這項理論,並認為原杉藻的學名Prototaxites(意為「第一個紫杉」[10])應該要更名為Nematophycus(意思為「絲狀的藻類」[11])[6],不過這項行為已嚴重違反了生物的命名規約,因此無法實行[12]。道森雖然不斷堅持自己的理論,但是原杉藻的結構在微觀上顯示他的理論是站不住腳的,在這之後道森也不斷提議將原杉藻改名(改成 Nematophyton,意思為「絲狀的植物」),意圖掩蓋自己當年將原杉藻誤認為是一種樹木的事實[6],但是基於國際藻類、真菌、植物命名法規,原杉藻的學名已確立並且無法更改,縱使意思與事實相違背,此學名仍然沿用至今。
雖然有許多證據顯示原杉藻生活於陸地上[13][14],卡魯瑟斯當時認為它們其實為一種海生的藻類,1919年,英格蘭植物學家阿圖·哈里·查爾赫則指出他不應排除其為真菌的可能性。由於缺乏任何可與現存生物進行比較研究的組織特徵,各種理論都難以獲得支持[6],因此原杉藻的分類地位的謎團遲遲未解。直到2001年,經過20年的研究,美國國立自然史博物館的Francis Hueber發表了一篇論文,提出證據表明原杉藻實屬一種真菌[6]。
這個理論最初受到了懷疑,但在之後出現了更多證據來支持這項理論[15]。2007年,由包括Hueber與芝加哥大學的C·凱文·博伊斯所組成的研究團隊針對原杉藻化石進行了同位素分析[1],結果亦支持原杉藻屬於真菌,因為原杉藻的化石中含有多種碳的同位素,而行光合作用的植物與藻類則僅會有少數幾種特定的碳同位素,這代表原杉藻的生存仰賴多種而非單一的碳來源,為異營生物(植物等自營生物多半僅有單一來源,也就是透過光合作用取得),和現代的真菌一樣以吸收基質中的各種有機物維生[1]。原杉藻可能有十分龐大的地下菌絲網路來吸收足夠的營養支持其生長,亦可能具有菌絲索而有能力進行長距離的營養輸送[16]。
關於原杉藻與各類群真菌的關係也有許多不同理論。Hueber認為原杉藻的化石中有子實層與原始擔子的痕跡,而認為其屬於擔子菌門[6]。也有觀點認為其應歸屬於球囊菌門或毛黴門[17]。2017年,有研究再次檢視原杉藻化石的構造,認為有子囊與側絲的痕跡,而主張其應歸屬於子囊菌門[3]。
有其他研究認為原杉藻可能是層層堆疊的地錢[18],但這項理論被認為可能性不大[19]。亦有其他研究主張原杉藻是是類似於紅豆杉屬的維管束植物、類似海帶目的藻類與具有類似真菌管狀組織的地錢門植物[18]。
與本屬相似的Nematasketum屬之化石組織中同樣也有類似的管束構造,代表它們可能也是真菌[20]。
古生態學
原杉藻可能為當時最高的陸生生物,當時最高的植物為頂囊蕨,高度僅有1(3英尺),其他陸地生物包括一些小型的無脊椎動物。維管束樹木及灌木的興起後原杉藻便滅絕了[4]。原杉藻高聳的結構可能可用來散播孢子,或用以競爭光線[4],芝加哥大學研究團隊則將原杉藻復原為無分枝的柱狀組織[21]。化石中發現與藻類相關的生物分子代表它們的表面可能被共生或寄生性的藻類所覆蓋,也可能其本身就是藻類[4][22][23]。
曾經有發現過入侵維管束植物的原杉藻菌絲化石[6],也曾有發現有動物居住於原杉藻其中的證據:動物挖掘造成的空洞遍布於原杉藻枝幹內,有理論認為這些動物的寄生可能導致原杉藻的滅絕[6]。然而在泥盆紀早期至晚期均有節肢動物在原杉藻枝幹內鑽孔的化石痕跡發現,這代表原杉藻的族群有能力承受這些動物所帶來的傷害達數百萬年[24]。之後植物才開始演化出木質化莖,這些動物也漸轉移到植物上鑽孔[24]。
參考文獻
- Boyce, K.C.; Hotton, C.L.; Fogel, M.L.; Cody, G.D.; Hazen, R.M.; Knoll, A.H.; Hueber, F.M. (PDF). Geology. May 2007, 35 (5): 399–402 [2019-03-26]. Bibcode:2007Geo....35..399B. doi:10.1130/G23384A.1. (原始内容存档 (PDF)于2011-09-28).
- Retallack G.J. & Landing, E. . Mycologia. 2014, 106: 1143–1156. PMID 24990121. doi:10.3852/13-390.
- Rosmarie Honegger, Dianne Edwards, Lindsey Axe, Christine Strullu-Derrien. . Philosophical Transactions of the Royal Society B. 2017, 373 (1739). doi:10.1098/rstb.2017.0146.
- Selosse, M.A. . Mycological Research. 2002, 106 (6): 641–644. doi:10.1017/S0953756202226313.
- Hillier, R; Edwards, D; Morrissey, L. B. . Palaeogeography, Palaeoclimatology, Palaeoecology. 2008, 270 (3–4): 366. doi:10.1016/j.palaeo.2008.01.038.
- Hueber, F.M. . Review of Palaeobotany and Palynology. 2001, 116 (1): 123–158. doi:10.1016/S0034-6667(01)00058-6.
- Schmid, Rudolf. . Science. 1976, 191 (4224): 287–288. Bibcode:1976Sci...191..287S. PMID 17832148. doi:10.1126/science.191.4224.287.
- Jonker, F.P. . Palaeontographica, B. 1979: 39–56.
- A fossil specimen collected by Charles Darwin's friend Joseph Dalton Hooker, was mislaid for 163 years at the British Geological Survey offices in London ("Scientists find lost Darwin fossils in gloomy corner of British Geological Survey", Christian Science Monitor, 17 January 2012 页面存档备份,存于; identifying Hooker as "John Hooker").
- The "Taxinaea" (Taxaceae) are the grouping of conifers to which Dawson drew analogy
- Almond Miller, Samuel. . author. 1877: 35.
- Seward, A. C. (Albert Charles), , University Press ; New York : Macmillan: 192, 1898 [13 February 2016], (原始内容存档于2018-08-01)
- Seward, A. C. . Cambridge University Press. 2010: 119. ISBN 9781108016001.
- William Dawson, Sir John. . Library of Alexandria. 2016. ISBN 9781465606853.
- Debra Lindsay (2005) Prototaxites Dawson, 1859 or Nematophycus Carruthers, 1872: Geologists V. Botanists in the Formative Period of the Science of Paleobotany. Earth Sciences History: 2005, Vol. 24, No. 1, pp. 35-61.
- Hillier RD, Edwards D, Morrissey LB. . Palaeogeography, Palaeoclimatology, Palaeoecology. 2008, 270: 366–380. doi:10.1016/j.palaeo.2008.01.038.
- G. J. Retallack, Ed Landing. . Mycologia. 2014, 106 (6). doi:10.3852/13-390.
- Graham, LE, Cook, ME, Hanson, DT, Pigg, KB and Graham, JM. . American Journal of Botany. 2010, 97 (2): 268–275. PMID 21622387. doi:10.3732/ajb.0900322.
- Taylor, T. N.; Taylor, E. L.; Decombeix, A. -L.; Schwendemann, A.; Serbet, R.; Escapa, I.; Krings, M. . American Journal of Botany. 2010, 97 (7): 1074. PMID 21616859. doi:10.3732/ajb.1000047.
- Edwards, D.; Axe, L. . Botanical Journal of the Linnean Society. 2012, 168 (1): 1–18. doi:10.1111/j.1095-8339.2011.01195.x.
- Prehistoric mystery organism verified as giant fungus 页面存档备份,存于 Press release from University of Chicago, April 23, 2007.
- Niklas, K.J. . Brittonia. 1976, 28 (1): 113–137. JSTOR 2805564. doi:10.2307/2805564.
- Niklas, K.J.; Pratt, L.M. . Science. 1980, 209 (4454): 396–7. Bibcode:1980Sci...209..396N. PMID 17747811. doi:10.1126/science.209.4454.396.
- Labandeira, C. . Insect Science. 2007, 14 (4): 259–275. doi:10.1111/j.1744-7917.2007.00152.x.
