囊泡藻类
囊泡藻界(学名:)是一類真核生物。囊泡藻界这个概念是湯瑪斯·卡弗利爾-史密斯1981年提出的色藻界的修订。表示双鞭毛生物与红藻发生单独的内共生后进化出的所有后代物种。 [1]囊泡藻界是2005年提出的真核生物域六个界之一。[2]
| 囊泡藻类 | |
|---|---|
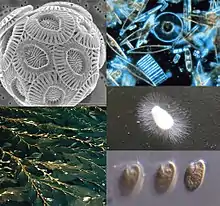 | |
| 左上起順時針:定鞭藻、矽藻、卵菌(Oomycota、水黴)、隱藻與褐藻(巨藻,Macrocystis)。 | |
| 科学分类 | |
| 总域: | 新壁總域 Neomura |
| 域: | 真核域 Eukaryota |
| 演化支: | 雙鞭毛生物 Bikonta |
| 演化支: | 囊泡藻类 Chromalveolata Cavalier-Smith, 1998 |
分类
历史上,许多囊泡藻界生物被认为是植物,因为它们具有细胞壁,具有光能合成的能力,很多情况下其形态类似于陆生植物. 但是当“五界说”(动物界、植物界、真菌界、原生生物界、原核生物界)流行起来之后,大多数囊泡藻类的生物都被分入原生生物界,水霉 (water mold) 与网粘菌 (labyrinthulids, or slime nets)被列入了真菌界,褐藻仍保持在植物界。
2005年,作为原生生物学的当时共识,囊泡藻界被列为真核生物的六个界之一[2]。不過尚未正式定為界層級。含以下四大分類群:
某些類群,例如纤毛虫与水黴,已经丧失了光能合成能力,但囊泡藻界生物大多数是自养。所有能光能合成的囊泡藻界生物使用叶绿素 a 与 c, 许多还使用辅助色素. 囊泡藻界生物使用类似的甘油醛-3-磷酸去氫酶蛋白质[6]。
早在2005年,就有研究质疑囊泡藻界是否是单系群[7] 。2006年,有评论论文认为真核生物六界系统中缺少很多证据[8]。 截至2012年,看来正在形成共识,把囊泡藻界分为两个类别。其中之一包括不等鞭毛门和囊泡虫总门,并把有孔虫界也列入,称之为SAR总群; 另一个类别包括了隐藻门与定鞭藻门[3][4]。 新近的研究把隐藻门与定鞭藻门分开,认为前者是SAR总群的姐妹群,后者应归入泛植物界[5]。
大量的名称用于囊泡藻界内部各类别的组合:
- Halvaria. 2007年到2008年的研究表明,不等鞭毛门和囊泡虫总门关系密切,因此把二者所在的进化支称为Halvaria.[3][4][9]
- SAR总群(SAR supergroup). 多孔虫界最初是与囊泡藻界并列。后来的研究表明多孔虫界应该与不等鞭毛门和囊泡虫总门有共同的起源。因此,多孔虫界加上Halvaria,称作SAR总群.[9][10]
- Hacrobia. 囊泡藻界中的另外两个类别,隐藻门与定鞭藻门,在某些分析中被认为是相近的[3][4],形成了一个进化支被称为Hacrobia。Hacrobia看来更接近泛植物界[3][4]。更早的时候,湯瑪斯·卡弗利爾-史密斯就已经建议把泛植物界与囊泡藻界合称为Corticata。如前述,新近的研究把隐藻门与定鞭藻门分开,认为前者是SAR总群的姐妹群,后者应归入泛植物界[5]。
形态
囊泡藻界没有非常普遍的共同形态特征。但其每个主要子类别都有独特的特征, 例如,囊泡虫总门具有的囊泡(alveoli), 定鞭藻门具有的附着鞭毛(haptonema), 隐藻门具有的喷射体(ejectisome),不等鞭毛门具有的两根不等长的鞭毛。但是这些特征都没有在囊泡藻界中普遍出现。
仅有的囊泡藻界的共同特征有:
- 有共同起源的色素体
- 大多数细胞壁都含有纤维素
对于这样的非常多样性的界,很难总结出其共同特性。
生态地位

许多囊泡藻界生物对生态系统有巨大的影响,往往是有害的影响。腰鞭毛虫门产生的赤潮会导致鱼类大量死亡,危害牡蛎的收获。顶复门是很多很成功的动物寄生虫. 卵菌纲的水霉会造成严重的植物病害. 实际上正是致病疫霉(Phytophthora infestans)引起了1845年-1852年的爱尔兰大饥荒.

但是,囊泡藻界的很多物种在生态系统中发挥了重要作用。硅藻是地球上重要的光合作用生物。褐藻产生了大量水下"森林"为海洋生物提供了栖息地与膳食。
囊泡藻界还提供了许多直接供人类使用的产品。褐藻胶用做食品增稠剂,在冰淇淋中常见。硅藻的硅质外壳有很多用处,如硅藻土用于过滤,用来做牙膏、做反光油漆。
参考文献
- Keeling PJ. . J. Eukaryot. Microbiol. 2009, 56 (1): 1–8. PMID 19335769. doi:10.1111/j.1550-7408.2008.00371.x.
- Adl, Sina M.; et al., , Journal of Eukaryotic Microbiology, 2005, 52 (5): 399 [2012-02-25], doi:10.1111/j.1550-7408.2005.00053.x, (原始内容存档于2017-09-14)
- Burki, Fabien; Shalchian-Tabrizi, Kamran & Pawlowski, Jan. . Biology Letters. 2008, 4 (4): 366–369. PMC 2610160. PMID 18522922. doi:10.1098/rsbl.2008.0224.
- Kim, E; Graham, LE. Redfield, Rosemary Jeanne , 编. (Free full text). PLoS ONE. Jul 2008, 3 (7): e2621 [2012-02-25]. PMC 2440802. PMID 18612431. doi:10.1371/journal.pone.0002621. (原始内容存档于2014-11-04).
- Burki, F.; Okamoto, N.; Pombert, J.F.; Keeling, P.J. . Proc. Biol. Sci. 2012. doi:10.1098/rspb.2011.2301.
- Takishita K, Yamaguchi H, Maruyama T, Inagaki Y. Zhang, Baohong , 编. . PLoS ONE. 2009, 4 (3): e4737 [2012-02-25]. PMC 2649427. PMID 19270733. doi:10.1371/journal.pone.0004737. (原始内容存档于2020-04-13).
- Harper, J. T., Waanders, E. & Keeling, P. J. 2005. On the monophyly of chromalveolates using a six-protein phylogeny of eukaryotes. Int. J. System. Evol. Microbiol., 55, 487-496. (PDF). [2010-04-26]. (原始内容 (PDF)存档于2008-12-17).
- Laura Wegener Parfrey, Erika Barbero, Elyse Lasser, Micah Dunthorn, Debashish Bhattacharya, David J Patterson, and Laura A Katz. . PLoS Genet. 2006 December, 2 (12): e220. PMC 1713255. PMID 17194223. doi:10.1371/journal.pgen.0020220.
- Fabien Burki, Kamran Shalchian-Tabrizi, Marianne Minge, Åsmund Skjæveland, Sergey I. Nikolaev, Kjetill S. Jakobsen, Jan Pawlowski. . PLoS ONE. 2007, 2 (8): e790 [2012-02-25]. PMC 1949142. PMID 17726520. doi:10.1371/journal.pone.0000790. (原始内容存档于2014-11-04).
- Hampl V, Hug L, Leigh JW; 等. . Proc. Natl. Acad. Sci. U.S.A. March 2009, 106 (10): 3859–64 [2012-02-25]. Bibcode:2009PNAS..106.3859H. PMC 2656170. PMID 19237557. doi:10.1073/pnas.0807880106. (原始内容存档于2020-04-10).