專性厭氧菌
專性厭氧菌(obligate anaerobes)是一類僅能進行無氧呼吸,且無法在正常大氣(氧含量21%)等富氧環境下存活的微生物[1][2]。這類生物對氧氣的耐受力不同,部分專性厭氧菌可以在氧含量達8%的環境下存活,但有些專性厭氧菌則僅能在氧含量低於0.5%的環境下存活[3]。值得注意的是,微需氧微生物雖然與專性厭氧菌一樣不能在正常大氣中存活(前者的生存環境中氧含量一般爲2%-10%),但它的呼吸方式是有氧呼吸,而不是專性厭氧菌的無氧呼吸或發酵[1][3][4]。
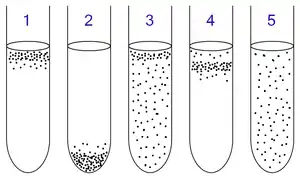
1中培養的是專性需氧微生物。因爲它們不能進行發酵及無氧呼吸,所以它們浮在了氧含量最高的上層。
2中培養的是專性厭氧菌。因爲它們體內因無過氧化氫酶等物質無法在有氧條件下存活,故浮在氧含量最低的下層。
3中培養的是兼性厭氧菌。它們既可以在有氧條件下也能夠在無氧條件下生存,所以它們分佈在試管培養基上的各處。但因爲有氧呼吸較無氧呼吸能產生更多的ATP(三磷酸腺苷),故更多的菌體分佈在上層。
4中培養的是微需氧微生物。它們不能進行發酵及無氧呼吸,也不能在高氧濃度環境中存活,故它們浮在試管中上層。
5中培養的是耐氧厭氧生物。這種細菌不能進行有氧呼吸,但卻可以在高氧濃度環境下存活,所以它們均勻地分佈在試管培養基上各處。
對氧的敏感性
專性厭氧菌之所以對氧敏感,是因爲:
- 三线态氧分子的外層分子軌道上有兩個未成對電子。這使得它容易在細胞內被還原爲對生物體有毒害作用的超氧化物(O−
2)或是過氧化氫(H
2O
2)[1]。好氧性生物體內含有過氧化氫酶和超氧化物歧化酶,它們可以將超氧化物和過氧化氫分解掉。但厭氧細菌體內卻很少甚至不含這些酶[1][2][3][5]。專性厭氧菌對氧氣的耐受能力(0.5%-8%)被認爲能夠反映其體內超氧化物歧化酶和過氧化氫分解酶的含量。[2][3] - 液體中溶解的氧氣會增加液體的還原電位,另外,高氧化還原電位會抑制某些專性厭氧菌的生長[3][5][6]比如說,產甲烷菌僅能在氧化還原電位低於-0.3 V的環境中生長[6]。
- 硫化物是一些酶的重要組成部分,而分子氧會氧化這些硫化物竝生成二硫鍵,造成部分酶(比如固氮酶)的活性降低。這些酶失活的話,專性厭氧菌可能就沒法正常存活了[1][5][6]。
- 專性厭氧細菌的生長可能會因爲電子全被用來還原氧造成的用作生物合成的還原當量下降而受到抑制[6]。
能量代謝
專性厌氧菌藉由無氧呼吸和發酵產生能量。在無氧呼吸中,葡萄糖先被轉化爲丙酮酸,而後者隨後又被轉化爲乙酰CoA。在有氧呼吸中,乙酰CoA隨後會經由電子傳遞鏈和三羧酸循環(檸檬酸循環)分解。無氧呼吸與有氧呼吸不同,一些電子受體(比如硫酸鹽、硝酸鹽、鐵、錳、汞、一氧化碳等)在它的電子傳輸鍊中取代了氧的位置[4]。
發酵與無氧呼吸不同,電子傳遞鏈沒有參與糖酵解生成的丙酮酸的分解(即沒有氧化磷酸化)。乳酸發酵、混合酸發酵、2-3丁二醇發酵均爲發酵的實例。[4]
在消耗等量有機物的情況下,無氧呼吸和發酵產生的能量(即ATP(三磷酸腺苷)的產量)比有氧呼吸要低[4]。這也是兼性厭氧菌在氧氣充足的條件下優先進行有氧呼吸的原因。兼性厭氧菌的此行爲可在它們被培養於巰基乙酸肉湯培養基時觀察到[1]。
例子

屬於專性厭氧菌的細菌包括放線菌、梭菌、大桿菌、梭桿菌、消化鏈球菌、普氏菌、丙酸桿菌和Veillonella菌等。與上述細菌不同,同屬專性厭氧菌的梭菌如破傷風梭菌及肉毒桿菌等在大氣氧濃度環境中能產生芽孢而呈休眠蟄伏[5],當芽孢到達適合生長的環境則開始生長並產生強力外毒素,造成人畜的疾病,詳見肉毒桿菌中毒。 大桿菌的炭疽菌形成的芽孢扺抗能力特強,唯有採用121°c,15-20分鐘的高壓減菌法方可殺滅,可隨空氣散布到各處,並蟄伏於土壤中,當人畜吸入芽孢或傷口接觸,則引發炭疽病。
參考
- Prescott LM, Harley JP, Klein DA. 3rd. Wm. C. Brown Publishers. 1996: 130–131. ISBN 0-697-29390-4.
- Brooks GF, Carroll KC, Butel JS, Morse SA. 24th. McGraw Hill. 2007: 307–312. ISBN 0-07-128735-3.
- Ryan KJ; Ray CG (editors). 4th. McGraw Hill. 2004: 309–326, 378–384. ISBN 0-8385-8529-9.
- Hogg, S. 1st. Wiley. 2005: 99–100, 118–148. ISBN 0-471-49754-1.
- Levinson, W. 11th. McGraw-Hill. 2010: 91–178. ISBN 978-0-07-174268-9.
- Kim BH, Gadd GM. . 2008.
- Carlile MJ, Watkinson, SC. . Academic Press. 1994: 33–34. ISBN 0-12-159960-4.
