米-门二氏动力学

方程与推导
以下米氏方程的推导是由Briggs和Haldane在1925年提出的[2]:
假设有下图所示的酶促反应

假设此酶促反应不可逆,反应产物不和酶结合;k2<k-1, E+S⇌ES 之间的平衡迅速建立达到平衡态(Steady-state),也就是受質和酶的化合物(ES)的浓度不变;建立平衡态所消耗的受質的量很小,可以忽略。这样有以下关系:


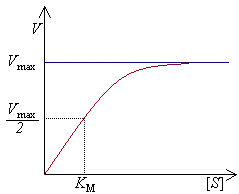
米氏常数Km的定义为:

原式可简化为:
(1)

总的酶的浓度[E0]等于自由酶[E]与酶-受質化合物[ES]的和,则有以下关系:

(2)

将(2)式代入(1):

整理得:


(3)

下式可以描述该酶促反应的速率:
(4)

将 (3) 代入 (4),分号上下同时乘以[S]得:
或

该式可通过非线性作图或Lineweaver-Burk(双倒数作图),Eadie-Hofstee等作图法变换为线性图进行分析。
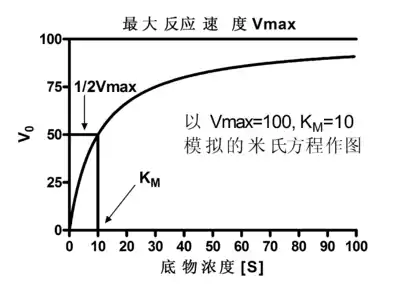
用Vmax=10,KM=10绘制的米氏方程的图像。
在推导过程中几点需要注意:
- [E0]是总的酶的量。反应中酶-底物配合物的量[ES]是极不好测量的,所以式子必须写成[E0]表示的形式,因为试验中所用的酶的量是已知的。
- d[P]/dt(V0, 反应初速度),是试验中测得的产物生成的初速度,一般是酶促反应在反应开始的几秒钟到几分钟之内的速度,在这段时间内底物的真实浓度几乎和底物最初的浓度相同([S]≈[S0])。
- k2[E0](Vmax) 是酶促反应在给定的酶的量下的最大速度(当所有的酶都在酶-底物配合物的状态下)。k2有时也写为kcat。
作用
- 米氏常数是酶的特征性物理常数。
- 从KM可判断酶的专一性和天然底物。
- 进行酶活力测定时通常选10KM。
- 底物浓度较低时,KM可判断底物走哪一条代谢途径。
试验方法
要测得方程中的KM和Vmax,需要在酶的量[E0]恒定并已知的情况下,在不同的底物浓度[S]下测得反应的初速度V0,用非线性作图或线性作图的方法求得KM和Vmax。
KM反映了底物和酶结合的紧密程度,Vmax反映了酶催化反应的速度。
参考资料
- Leonor Michaelis, Maud Menten (1913). Die Kinetik der Invertinwirkung, Biochem. Z. 49:333-369.
- G. E. Briggs and J. B. S. Haldane (1925) A note on the kinetics of enzyme action, Biochem. J., 19, 339-339.
This article is issued from Wikipedia. The text is licensed under Creative Commons - Attribution - Sharealike. Additional terms may apply for the media files.
