分支龍屬
分支龍屬(屬名:),又名歧龍、阿利奧拉龍,是獸腳亞目暴龍科下的一個屬,生活於上白堊紀的蒙古。分支龍是由蘇聯古生物學家Sergei Kurzanov在1976年所命名。模式種是遙遠分支龍(A. remotus),化石包含一個部份顱骨、三個蹠骨,發現於蒙古,該地在7000萬年前到6500萬年前是片氾濫平原。第二個種是阿爾泰分支龍,化石是一個更完整的骨骼,是由Stephen L. Brusatte等人在2009年命名。
| 分支龙属 化石时期:白堊紀晚期,70 Ma | |
|---|---|
 | |
| 分支龍的復原圖 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龙总目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 兽脚亚目 Theropoda |
| 科: | †暴龍科 Tyrannosauridae |
| 亚科: | †暴龍亚科 Tyrannosaurinae |
| 族: | †分支龙族 Alioramini |
| 属: | †分支龙属 Alioramus Kurzanov, 1976 |
| 模式种 | |
| 遙遠分支龍 Alioramus remotus Kurzanov, 1976 | |
| 種 | |
| |
如同大部分獸腳類恐龍,分支龍是二足恐龍,而銳利的牙齒顯示牠們是肉食性動物。牠比近親的暴龍及特暴龍較小型,但目前僅有發現兩個標本,而且可能來自於幼年體或亞成年體,因此難以估計成年個體的大小。分支龍的特徵是在其口鼻部上側的一排五個骨冠,以及牙齒較其他暴龍科更多。分支龍的頭顱骨較其他暴龍科為低矮,這可能是個幼年特徵。
分支龍與其他暴龍科動物的關係未確定,但部分科學家認為分支龍與生存於同時代、同地區的特暴龍是近親,或者分支龍是特暴龍的幼年個體。根據新發現的阿爾泰分支龍的化石,分支龍應該是特暴龍的近親。
敘述

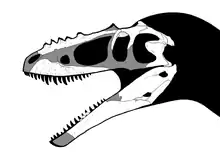

根據Sergei Kurzanov在1976年的最初研究,分支龍的身長估計是在5到6公尺之間[1]。但是顱骨在化石化的過程中遭到變形,這意味者該個體的身長較Kurzanov的估計值更短。另外,如果這個標本是個幼年體,成年分支龍的體型應該更大,但目前還沒有發現成年體的化石[2]。
分支龍的頭顱骨約為45公分長[3],頭顱骨長而低矮,外形類似原始暴龍超科,以及幼年暴龍科的頭顱骨。遙遠分支龍的前上頜骨部份沒被發現,但頭顱骨的高度大於寬度,與已知所有的暴龍超科相同[2]。鼻骨互相癒合,上有一排骨質瘤,沿者鼻骨接合處排列,共五個。這些骨質瘤的高度大於一公分[1]。很多暴龍超科恐龍都有凹凸不平的鼻部,但有骨質瘤的就有分支龍及阿巴拉契亞龍(有六個)[4]。
頭顱骨後部有個橫向突出物,從癒合頂骨延伸出來,稱為頸脊(Nuchal crest),這是所有暴龍超科都有的特徵。分支龍的頸脊明顯較厚,類似特暴龍與暴龍。如同頭顱骨其他部分,分支龍的下頜修長,可能是個幼年個體的特徵[2]。分支龍的下頜隅骨側邊稜脊,連接者齒骨後方的方形突,使這兩塊骨頭固定者,減少下頜的靈活度,在暴龍科之內,只有分支龍與特暴龍具有這個特徵[5]。分支龍亦有另一個特徵,就是在上頜及下頜都有很多牙齒。其他暴龍科的前上頜骨牙齒兩邊各有4顆,牙齒橫剖面為D字型。分支龍的上頜骨各有16隻牙顆,而在齒骨則各有18顆,比已知的暴龍科更多[6]。阿爾泰分支龍的腦殼特徵,介於原始獸腳類恐龍、鳥翼類之間[7]。
遙遠分支龍的顱後身體部份僅發現三塊蹠骨。阿爾泰分支龍的化石更為完整,使科學家能更清楚分支龍的外貌型態[8]。但從已知暴龍科來推測,分支龍應該與以二足行走;而後肢比例極可能類似似鳥龍下目,脛骨與蹠骨長,兩者總長約等於股骨。分支龍的前肢應該很小,上有兩指,不過某些暴龍科標本上有退化的第三指。長尾巴可平衡頭部與身體的重量,將重心維持在臀部位置[2]。

發現與命名
分支龍的正模標本(編號PIN 3141/1)是一個部份騰顱骨,與三個相連的蹠骨;是在70年代早期,由一個蘇聯蒙古的挖掘團隊在蒙古的巴彥洪戈爾省Nogon-Tsav地層中挖掘出土,並由俄羅斯古生物學家Sergei Kurzanov於1976年所命名。牠的冠狀物、口鼻部較長、與低矮頭顱骨,都與其他暴龍亞科有所不同,所以被命名Kurzanov為遙遠分支龍或偏僻分支龍(A. remotus),屬名由拉丁文的「alias」(即「其他」的意思)及「ramus」(即「支部」的意思)構成,意指牠們是另一個全新及不同的暴龍亞科分類單元;同樣地,種名「remotus」在拉丁文是「遙遠」的意思,意指牠們的差別性[1]。目前只有發現兩個標本,分別屬於遙遠分支龍、阿爾泰分支龍[2]。
分類
分支龍被古生物學家分類於暴龍超科之中,但過去因為其化石的資訊有限,因此不能做出更精準的分類。新標本的發現,使分支龍的分類與演化關係更為清楚[2]。在2003年,菲力·柯爾(Phil Currie)提出分支龍與暴龍、特暴龍、懼龍,共同屬於暴龍亞科[4]。在2004年,湯瑪斯·荷茲(Thomas Holtz)則認為分支龍有可能屬於暴龍亞科,但也有相同的可能性是不屬於暴龍科的,原因是牠們的可能幼年體特徵,也許是基礎暴龍超科的特徵[2]。在2005年,另一個研究則因為牠們化石的破碎性,而不將分支龍包含在內[9]。在2009年,新發現的阿爾泰分支龍化石,確定分支龍屬於暴龍亞科[8]。
分支龍與特暴龍生存於相同時代的相同地區,兩者具有許多相同的頭部特徵,例如:下頜的齒骨與隅骨間癒合、鼻骨與淚骨間缺乏骨質的連結,後者是暴龍科中除了分支龍、特暴龍、懼龍以外成員都有的特徵。因此分支龍與特暴龍可能是近親,兩者構成暴龍科的亞洲分支[4][5]。在特暴龍的某些標本上,鼻部上有一排骨質瘤,類似分支龍,但較為低矮。遙遠分支龍的顱骨長而低矮,顯示該個體在死亡時尚未成熟,可能是特暴龍的幼年個體。但是,分支龍的更明顯骨質瘤與較多的牙齒,證明牠們與特暴龍是不同的物種[6]。阿爾泰分支龍的發現,更確定分支龍與特暴龍的獨立的屬[8]。標本更證明特暴龍的幼年與成年個體的牙齒數目一樣[10][11]。
古生物學
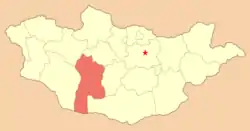
分支龍的化石標本是在蒙古的巴彥洪戈爾省Nogon-Tsav地層被發現,該地層被認為與耐梅蓋特組年代相同[1]。該地層沒有經過放射性定年法測定,該化石紀錄中的動物群顯示,該地層的年代是上白堊紀的麥斯特里希特階[12],距今約7400萬道6500萬年前[13]。
蒙古的麥斯特里希特階地層有耐梅蓋特組與Nogon-Tsav地層,兩者的特徵是潮濕的氣候,而其下層的巴魯恩戈約特組與德加多克塔組年代較早,氣候為半乾旱。耐梅蓋特組的沉積層為氾濫平原、大型河道、與古代土壤,而硝石沉積層顯示該地有週期性的旱季[14]。與較早期的地層相比,耐梅蓋特組有更多樣性、更大量的恐龍動物群。Kurzanov還發現了其他獸腳類,例如:特暴龍、似鳥龍下目、鐮刀龍超科[1],但沒有詳細的研究。在耐梅蓋特組,掠食動物可能以蜥腳類的泰坦巨龍類恐龍為食[5]。Nogon-Tsav地層並沒有大量發現其他恐龍化石,如果該地的動物群與耐梅蓋特組相似,應也會發現傷齒龍科、厚頭龍下目、甲龍科、鴨嘴龍類等化石[12]。泰坦巨龍類也是耐梅蓋特組的掠食動物的可能獵物[5]。
參考資料
- (俄文)Kurzanov, Sergei M. . The Joint Soviet-Mongolian Paleontological Expedition Transactions: 93–104.
- (英文)Holtz, Thomas R. . Weishampel, David B.; Dodson, Peter; & Osmólska, Halszka (eds.) (编). Second Edition. Berkeley: University of California Press. 2004: 111–136. ISBN 0-520-24209-2.
- (英文)Currie, Philip J. . . Cambridge: Cambridge University Press. 2000: 434–455. ISBN 978-0521545822.
- (英文)Currie, Philip J.; Hurum, Jørn H; & Sabath, Karol. (PDF). Acta Palaeontologica Polonica. 2003, 48 (2): 227–234 [2009-12-03]. (原始内容存档 (PDF)于2018-08-19).
- (英文)Hurum, Jørn H.; & Sabath, Karol. (PDF). Acta Palaeontologica Polonica. 2003, 48 (2): 161–190 [2009-12-03]. (原始内容存档 (PDF)于2018-09-21).
- (英文)Currie, Philip J. (PDF). Acta Palaeontologica Polonica. 2003, 48 (2): 191–226 [2009-12-03]. (原始内容存档 (PDF)于2018-08-19).
- (英文). PLOS Collections. [2012-11-09]. (原始内容存档于2012-04-01).
- (英文)Brusatte, Stephen L.; Carr, Thomas D.; Erickson, Gregory M.; Bever, Gabe S.; and Norell, Mark A. . Proceedings of the National Academy of Sciences of the United States of America. 2009,. online preprint (41): 17261–6. PMC 2765207. PMID 19805035. doi:10.1073/pnas.0906911106.
- (英文)Carr, Thomas D.; Williamson, Thomas E.; & Schwimmer, David R. . Journal of Vertebrate Paleontology. 2005, 25 (1): 119–143 [2008-02-01]. (原始内容存档于2020-07-27).
- (俄文)Maleev, Evgeny A. . Doklady, Academy of Sciences USSR. 1955, 104 (5): 779–783.
- (英文)Currie, Philip J. . Canadian Journal of Earth Sciences. 2003, 40 (4): 651–665. doi:10.1139/e02-083.
- (英文)Jerzykiewicz, Tomasz; & Russell, Dale A. . Cretaceous Research. 1991, 12 (4): 345–377. doi:10.1016/0195-6671(91)90015-5.
- (英文)Gradstein, Felix M.; Ogg, James G.; & Smith, Alan G. . Cambridge: Cambridge University Press. 2005: 500pp. ISBN 978-0521781428.
- (英文)Osmólska, Halszka. . Currie, Philip J.; & Kevin Padian (eds.) (编). . San Diego: Academic Press. 1997: 471–472. ISBN 0-12-226810-5.

{kind=link}