太古角龍屬
太古角龍(屬名:Yehuecauhceratops,意為「古代的有角臉」)是一屬尖角龍亞科的角龍科恐龍,來自白堊紀晚期的墨西哥科阿韋拉。屬下包含單一物種:模式種沙博館太古角龍(Yehuecauhceratops mudei)由Rivera-Sylva等人於2016年根據兩個部分骨骼進行敘述[1],並於2017年正式命名。[2]牠是小型的尖角龍亞科,身長只有3公尺,小於棲息同地區的另外兩種角龍科:阿古哈角龍和科阿韋拉角龍;三者可能形成棲位適應上的分化。鱗狀骨基部附近發育一道帶有單一粗糙突起的稜脊,可能支撐著一個小角,使太古角龍能從其他尖角龍亞科中鑑別出來。根據形態上的相似性,太古角龍與大鼻角龍、愛氏角龍關聯在一起,牠們一起構成大鼻角龍族演化支。[1][2]
| 太古角龙属 化石时期:坎潘階,72 Ma | |
|---|---|
 | |
| 骨架模型,展示於墨西哥科阿韋拉州薩爾蒂約的沙漠博物館 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龙总目 Dinosauria |
| 目: | †鸟臀目 Ornithischia |
| 科: | †角龙科 Ceratopsidae |
| 亚科: | †尖角龍亞科 Centrosaurinae |
| 族: | †大鼻角龙族 Nasutoceratopsini |
| 属: | †太古角龙属 Yehuecauhceratops Rivera-Sylva et al., 2017 |
| 模式种 | |
| 沙博館太古角龍 Yehuecauhceratops mudei Rivera-Sylva et al., 2017 | |
發現及命名

太古角龍的標本是於2007年至2011年的挖掘活動中,在德州大彎曲國家公園以南23.3公里,墨西哥科阿韋拉州奧坎波市薩拉達鎮(La Salada)附近地點所發現。[2]該地點暴露出的岩層屬於阿古哈組(Aguja Formation),雖然其位於層位垂直範圍的精確位置難以判定,但估計是坎潘階。[1]標本目前存放於沙漠博物館(Museo del Desierto)的古生物館藏中。[2]
正模標本CPC 274是一個部分骨骼,包含一個幾乎完整的左鱗狀骨、三個頂骨碎片、一個來自前上頜骨和齒骨的碎片、一個完整的肩胛骨和股骨、髖臼前的部分髂骨、以及一個神經棘斷裂的部分背椎。[1]其他所知骨骼還有部分肋骨、支持尾部的肌腱碎片、以及雜亂的碎片。[2]另外還有一個脛骨碎片,對於CPC 274標本來說太小了,這個標本被編入CPC 1478,2017年的研究並未提及。[1]
雖然這兩個標本於2016年由Rivera-Sylva等人敘述成尖角龍亞科的未定物種,牠們於2017年根據鱗狀骨中的鑑定特徵被正式命名成新屬。屬名Yehuecauhceratops由納瓦特爾語的yehuecauh(古代)加上希臘文字根ceratops(有角的臉)所組成;種名mudei取自沙漠博物館Museo del Desierto的縮寫MUDE。[2]
敘述
太古角龍是種小型角龍科,身長約3公尺。[2]正模標本根據脊椎縫線癒合狀況來推斷,可能是個晚期亞成體或成年體。[1][3]
頭骨材料

鱗狀骨是唯一可以將太古角龍從其他尖角龍亞科鑑別出來的材料。其與頂骨間的邊界有一梯狀邊緣,是尖角龍亞科的典型特徵。鱗狀骨長邊長於寬邊[1],不像許多進階尖角龍亞科(如戟龍、尖角龍、厚鼻龍[4]),但與愛氏角龍較類似[5]。鱗狀骨延伸的遠端部分長度為骨頭其餘部分的兩倍。[2]
沿著鱗狀骨外緣,有三個突出的波紋,可能支撐著褶飾(epiossification),但褶飾本身未保存下來。[2]第二條波紋略微向上偏轉,如同進階尖角龍亞科。[4][6][7]然而在溫氏角龍[8]和其他更進階的尖角龍亞科中,有至少四到五條波紋,而非只有三個。鱗狀骨底部附近發育一道稜脊,但不如大鼻角龍的突出,[9]且沒有延伸到鱗狀骨的扇形背側。不像愛氏角龍、溫氏角龍[8]、以及亞伯達角龍[6],太古角龍缺乏沿著稜脊的三道平滑突起;[1]只在稜脊中央有一個粗糙突起,可能長著一支小角。稜脊位置、單一突起這兩項特徵為太古角龍的自衍徵,使牠能夠區別於其他尖角龍亞科。[2][1]

頂骨相對光滑。最大和最厚的碎片來自頸盾中心附近,代表太古角龍可能有相對較小的孔,不同於其他尖角龍亞科,而類似愛氏角龍。[5]一個頂骨褶飾保存下來;具簡單的月牙形邊緣,形狀不像鱗狀骨的波紋,且表面粗糙。如同惡魔角龍、愛氏角龍、異角龍、大鼻角龍,這個褶飾不和其他褶飾直接相鄰或重疊,反而在褶飾周圍的骨頭有向內的起伏。[1]
保存下來的部分前上頜骨屬於鼻孔邊緣,一道沿骨頭斜向延伸的稜脊類似惡魔角龍,[10]並比愛氏角龍的更突出。[5]圍繞鼻孔的頂部以及前上頜骨內表面都粗糙化,鼻孔也是骨頭的最厚的部分。根據下頜喙突來判斷,保存下來的部分齒骨可能來自下頜的後部三分之一。這保存了一個齒列,其中有六個齒槽長出三個牙齒,牙齒由骨頭中心向動物的中線偏移。[1]
顱後材料
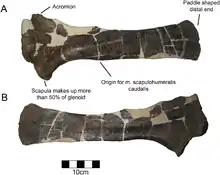
如同典型的尖角龍亞科,肩胛骨構成關節盂的一半。如同其他角龍科,肩峰突出且非常粗糙。骨頭底緣的最厚處陡峭角度為25度而非逐漸彎曲。肩胛骨頂端變寬成槳狀,比其他尖角龍亞科突出。[4]如同其他角龍科,髂骨大幅偏轉,頂視呈筆直的。[1]
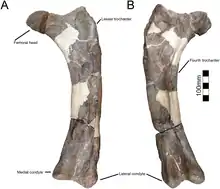
強健的股骨呈桿狀、筆直幾乎不彎曲,如同愛氏角龍,[5]雖然可能是因為化石受到壓縮。股骨中,股骨頭圓潤呈球形,大轉子位置高於股骨頭之上,而第四轉子形成一個突出的冠,如同其他尖角龍亞科。脛骨的脛脊擴張,約是擴張端的40%寬,類似愛氏角龍。[5][4][1]
一個背椎,根據大神經管判斷,可能來自靠近頸部至軀幹間的過渡帶,[11]邊緣略縮,於神經管兩側各有一凹陷。脊椎關節兩端均圓形凹入(雙內凹)。神經管高度長於寬度的25%。包圍神經管的神經弓幾乎完全融合進骨頭的其餘部分。但在某處可以觀察到少量縫線。[1]
分類

2016年的系統發生學分析發現太古角龍位於尖角龍亞科一個非常基礎的位置,並與愛氏角龍、大鼻角龍共享數個特徵,因此屬於同一演化支,這個類群後來被命名成大鼻角龍族。詳細演化樹如下所示:[1]
| 尖角龍亞科 Centrosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
太古角龍獨特、粗糙的鱗狀骨突起特徵只在NMMNH P-25052標本(一個來自新墨西哥州、較古老的米尼菲組的頭骨)以及來自亞歷桑納州、較古老的克里騰登堡組的克里滕登角龍屬也出現。[9][12]這些標本時間相距超過100萬年,不太可能是同一物種。此外UMNH VP16704標本(暱稱為"乳頭丘頭骨")的鱗狀骨扇形末端也類似太古角龍和NMMNH P-25052標本,但缺乏這種奇異的突起。[1]
古生態學
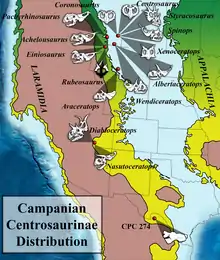
太古角龍來自阿古哈組(Aguja Formation)下部層位,當時這裡的生活環境是一個河口三角洲,由淡水至鹹水交界處的沿海沼澤和氾濫平原所構成。該地沉積物還包含牛軛湖痕跡、沿海海相沉積。雖然阿古哈組墨西哥地區的年代尚未確認,但根據同屬阿古哈組的其他地區,以及與其對應的普韋布洛組(Cerro del Pueblo Formation)的礦床測年來推斷,地質年代大致落在7250萬年前左右。[1]在墨西哥,阿古哈組的物種多樣性受到某種程度上的限制。除了太古角龍,當地其他發現包含開角龍亞科的阿古哈角龍;厚頭龍類的牙齒;來自鴨嘴龍科的巨大脛骨;鱷魚牙齒;龜鱉殼碎片。[1]
阿古哈組所對應的德州地區則有數種已知的恐龍:阿古哈角龍、櫛龍亞科的小貴族龍、厚頭龍科的德克薩斯頭龍、以及根據牙齒建立的屬理察伊斯特斯龍、未命名的賴氏龍亞科、結節龍科、暴龍科、似鳥龍科、近頜龍科、奔龍科。[13]非恐龍物種則有:大型鱷類如恐鱷[13][14]、稜角鱗鱷科;有鱗目的Odaxosaurus、Proxestops、Restes、Sauriscus、和其他多個未命名物種;烏龜屬Baena和Aspideretes,還有別的龜鱉目;滑體亞綱的Albanerpeton、蠑螈和青蛙;哺乳類的Cimolomys、Meniscoessus、Cimolodon、Alphadon、Turgidodon、Pediomys、Gallolestes等;多種硬骨魚;鮫魟類的弓鮫、滑齒鯊屬、Scapanorhynchys、帆鋸鰩屬、Ischyrhiza、Squatirhina、Ptychotrygon等。[14]
雖然角龍科在科羅拉多州北部的拉臘米迪亞沉積層很常見,但牠們在南方卻很罕見,而實際上保存下來的化石證據更少。雖然尖角龍亞科的化石材料有限,牠們某種程度上在南方更為常見;[1]然而墨西哥也得知了許多種開角龍亞科,如阿古哈角龍和科阿韋拉角龍(普韋布洛組)。這樣的現象與坎潘階拉臘米迪亞南北部受氣候影響的植物相差異相呼應;尤其是在德州的阿古哈組中,雙子葉植物的Metcalfeoxylon和Agujoxylon占主要優勢;[15]相較之下,北方的針葉樹較為豐富。在阿古哈組中,科阿韋拉角龍最大型,而太古角龍相對較小型,阿古哈角龍介於兩者之間,這可能代表著這三種角龍科具有不同程度影響體型的覓食習性。[2]
參考資料
- Rivera-Sylva, H.E.; Hendrick, B.P.; Dodson, P. . PLOS One. 2016, 11 (4): e0150529. PMC 4830452. PMID 27073969. doi:10.1371/journal.pone.0150529.
- Rivera-Sylva, H.E.; Frey, E.; Stinnesbeck, W.; Guzman-Gutirrez, J.R.; Gonzalez-Gonzalez. . Journal of South American Earth Sciences. 2017, 75: 66–73. doi:10.1016/j.jsames.2017.01.008.
- Brochu, C.A. . Journal of Vertebrate Paleontology. 1996, 16 (1): 49–62. doi:10.1080/02724634.1996.10011283.
- Ryan, M.J.; Holmes, R.; Russell, A.P. . Journal of Vertebrate Paleontology. 2007, 27 (4): 944–962. doi:10.1671/0272-4634(2007)27[944:AROTLC]2.0.CO;2.
- Penkalski, P.; Dodson, P. . Journal of Vertebrate Paleontology. 1999, 19 (4): 692–711. doi:10.1080/02724634.1999.10011182.
- Ryan, M.J. . Journal of Paleontology. 2007, 81 (2): 376–396. doi:10.1666/0022-3360(2007)81[376:ANBCCF]2.0.CO;2.
- Farke, A.A.; Ryan, M.J.; Barrett, P.M.; Tanke, D.H.; Braman, D.R.; Loewen, M.A.; Graham, M.R. . Acta Palaeontologica Polonica. 2011, 56 (4): 691–702. doi:10.4202/app.2010.0121.
- Evans, D.C.; Ryan, M.J. . PLOS One. 2015, 10 (7): e0130007. PMC 4496092. PMID 26154293. doi:10.1371/journal.pone.0130007.
- Sampson, S.D.; Lund, E.K.; Loewen, M.A.; Farke, A.A.; Clayton, K.E. . Proceedings of the Royal Society B. 2013, 280 (1766): 20131186. PMC 3730592. PMID 23864598. doi:10.1098/rspb.2013.1186.
- Kirkland, J.I.; DeBlieux, D.D. . Ryan, M.J.; Chinnery-Algeier, B.; Eberth, D.A. (编). . Indiana University Press. 2010: 117–140. ISBN 978-0-253-00779-7. JSTOR j.ctt16gzgng.
- Giffin, E.B. . Paleobiology. 1990, 16 (4): 448–458. doi:10.1017/S0094837300010186.
- Williamson, T.E. . Lucas, S.G.; Estep, J.W.; Williamson, T.E.; Morgan, G.S. (编). . New Mexico Museum of Natural History Bulletin 11. 1997: 51–59.
- Longrich, N.R.; Sankey, J.; Tanke, D. . Cretaceous Research. 2010, 31 (2): 274–284. doi:10.1016/j.cretres.2009.12.002.
- Rowe, T.; Cifelli, R.L.; Lehman, T.H.; Weil, A. . Journal of Vertebrate Paleontology. 1992, 12 (4): 472–493. JSTOR 4523473. doi:10.1080/02724634.1992.10011475.
- Lehman, T.M.; Wheeler, E.A. . PALAIOS. 2001, 16 (1): 102. doi:10.1669/0883-1351(2001)016<0102:AFDWFF>2.0.CO;2.
