尾綜骨
尾綜骨(英語:)是鳥類部分尾椎癒而形成的結構,它用於支撐尾部羽毛和肌肉。尾綜骨是尾臀(英語:)的主要組成部分,這一隆起可以在被烹飪的鳥類的後面清晰看見(如雞和火雞)
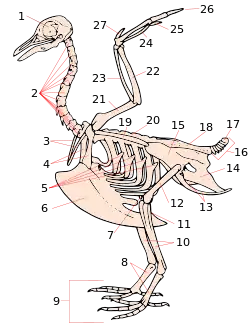
鴿子骨架,尾綜骨的編號為17
_(19178230020).jpg.webp)
北方鷂的軀幹骨骼圖繪(腹面朝右),脊柱末端的鐮刀形骨骼即為尾綜骨
進化
在1億3000至4000萬年的白堊紀早期,這一結構就開始演化。已知最早具有該結構的物種是孔子鳥。這種結構具有進化優勢,因為在始祖鳥等物種中發現的完全可移動的尾翼不利於其控制飛行。 現代鳥類在其胚胎狀態下仍會發育較長的尾椎骨,後來融合形成尾綜骨。 尾綜骨有兩種主要類型:第一種見於孔子鳥、反鳥類和其他中生代鳥類,以及像竊蛋龍科的天青石龍,牠們的尾綜骨長而呈桿狀或匕首狀。 具有這種尾綜骨的恐龍均未顯示出發達的舵羽。這些動物的尾羽由柔軟的絨毛組成,有時由2至4個中央“飄帶”組成,例如在孔子鳥的一些標本或副原鳥中發現的那樣[1]。 相比之下,尾綜骨在陸地天青石龍中的功能尚不清楚。但是,值得注意的是,它的年長親戚尾羽龍沒有尾羽,而具有對的稱羽毛,這些羽毛可能用於社交場合[2]。也許這樣的裝飾品在近頜龍超科及其親緣物種中很普遍,竊蛋龍科的動物最終進化出了尾綜骨來支撐它們。 另一種尾綜骨類型是犁頭形的。 它在扇尾類(鳥類及其近親)中發現,幾乎在所有飛行物種中都與操縱中使用的一系列發達的舵羽。 它們的中央直接與尾綜骨相連,就像在孔子鳥中一樣。 扇尾類的其他骨架被固定在適當的位置,並通過稱為舵羽球(拉丁語:,英語:)的結構移動。 已知最古老具有這種方式的物種是Hongshanornis longicresta[3]。[4]
參考資料
- Clarke et al. (2006)
- Xu, X.; Zheng, X.; You, H. . Nature. 2010, 464 (7293): 1338–1341. PMID 20428169. doi:10.1038/nature08965.
- O'Connor, J.K.; Gao, K.-Q.; Chiappe, L.M. (PDF). Journal of Vertebrate Paleontology. 2010, 30 (2): 311–321. doi:10.1080/02724631003617498.
- Clarke, Julia A.; Zhou, Zhonghe; Zhang, Fucheng. . Journal of Anatomy. 2006, 208 (3): 287–308. PMC 2100246. PMID 16533313. doi:10.1111/j.1469-7580.2006.00534.x.
外部链接
This article is issued from Wikipedia. The text is licensed under Creative Commons - Attribution - Sharealike. Additional terms may apply for the media files.
