形态发生
形态发生(希臘語:,字面意思是"形状的开始")是一个生物体发展形态的生物过程。它和细胞生长之調控、细胞分化被視為发育生物学的三个基本主題之一。
这个过程控制了细胞在胚胎发育过程中的空间分布。形态发生也可以发生在一个成熟的生物体,在细胞培养或肿瘤细胞群中。形态发生也描述了单细胞生命的发展形式,这些生命形式在它们的生命周期中没有胚胎阶段,也可以描述一个分类群体内一个身体结构的演化过程。
形态发生反应可能是由荷尔蒙、环境化学物质引起的,例如其他生物体产生的物质,作为污染物释放的有毒化学物质或放射性核素,以及其他植物,或由细胞空间图案引起的应激性。
历史
关于物理过程和限制如何影响生物生长的一些最早的想法和数学描述,例如一些自然模式,如某些生物的螺旋形,是由达西・汤普森在他1917年的著作《生长与形式》中提出的,另一些则是由阿兰·图灵在他的《形态发生的化学基础》(1952)中所描写。[1][2][3]汤普森解释了动物身体的形状是由不同方向不同的生长速度造成的,例如,蜗牛的螺旋壳的原理。图灵则正确地预测了一种形态发生的机制,两种不同的化学信号的扩散,一个激活和一个失活的生长,以及建立发展模式,这些成果都是在科学界观察到这种模式真正成因的几十年前被研究出来的。[4]对于实际生物体发展所涉及的机制的更充分了解,需要等到在1953年发现DNA结构,以及分子生物学和生物化学的发展之后了。
基因和分子基础
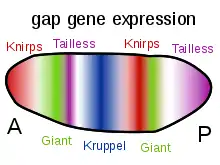
几种类型的分子在形态形成中是很重要的。形态物是可溶性分子,可以通过浓度梯度传播和携带控制细胞分化的信号。形态细胞通常通过与特定的蛋白受体结合而发挥作用。参与形态发生的一类重要分子是转录因子蛋白,通过与DNA的相互作用来决定细胞的命运。这些基因可以通过主管调控基因进行编码,并激活或破坏其他基因的转录;反过来,这些次级基因产物可以调节在基因调控网络的调控级联中的其他基因的表达。在这个级联的末端是控制细胞迁移等细胞行为的分子类,或者更广泛地说,它们的特性,如细胞粘附或细胞收缩。例如,在调节过程中,干细胞簇脱离细胞与细胞的粘附,成为迁移,在胚胎中重新激活特定细胞粘附蛋白并形成新的组织和器官。发育信号通路涉及形态发生包括Wnt、Hedgehog和ephrins。[5]
细胞基础
在组织层面上,无视控制手段,形态发生是由于细胞生长和运动。[6]形态发生也涉及细胞结构的变化或细胞在组织中的相互作用。这些变化可能导致组织的伸长、变薄、折叠、入侵或分离成不同的层。后一种情况通常称为细胞分类。细胞"排序"包括移动的单元格,以便排序成最大化同类型细胞之间接触的簇。由于马尔科姆·斯坦伯格通过差异性粘附假说提出了细胞粘附的能力。组织分离也可以通过更戏剧性的细胞分化事件发生,在这些事件中上皮细胞变成了间充质(见上皮-间充质过渡)。由于细胞粘附性和收缩性能的变化,间充质细胞通常会离开上皮组织。 在上皮-间充质转变后,细胞可以从上皮细胞迁移出去,然后与其他类似细胞在一个新的位置相关联。[7]
细胞-细胞粘附
在胚胎发育过程中,由于不同的亲缘关系,细胞被限制在不同的层面。其中一种方法就是细胞分享相同的细胞间粘附分子。例如,同源细胞粘附能够保持具有不同粘附分子的细胞群之间的边界。此外,细胞可以根据细胞间粘附的差异进行分类,所以即使是两个具有不同水平的相同粘附分子的细胞也可以分离出来。在细胞培养细胞中,具有最强的粘附力的细胞聚集体的中心。 而且,细胞与细胞间的粘附通常是通过细胞收缩来调节的,这会对细胞-细胞接触产生作用,从而使两个具有同等粘附分子水平的细胞可以分离出来。 负责粘附的分子称为细胞粘附分子(CAMs)。 已知的细胞粘附分子有几种类型,其中一种主要的分子是钙粘蛋白。在不同的细胞类型中,有几十种不同的地籍。钙粘附素以类似的方式与其他钙粘蛋白结合:E-cadherin(在许多上皮细胞上发现)优先结合其他的钙粘附分子。 间充质细胞通常表达其他类型的钙粘附素,如N-cadherin。[8][9]
细胞外间质
细胞外间质(ECM)负责将组织分离,提供结构性支持或提供细胞迁移的结构。胶原蛋白、层粘连蛋白和纤维连接素是主要的ECM分子,它们分泌并组装成薄片、纤维和凝胶。被称为整合素的多亚单位跨膜受体被用来与ECM结合。整合素在细胞外结合纤维连接素、层粘连蛋白或其他ECM成分,并在细胞内结合微丝结合蛋白质-酰胺和滑石蛋白,使细胞骨架与外部细胞结合。整合素在与ECM结合时也作为触发信号转导级联的受体。一个经过深入研究的包括乳腺导管分支的形态发生的例子。[10][11]
细胞收缩
组织可以通过细胞收缩来改变它们的形状并分成不同的层。就像肌細胞一样,肌球蛋白可以收缩细胞质的不同部位来改变其形状或结构。在胚胎组织形态发生过程中,我们可以看到肌球蛋白在胚胎组织形态发生中的收缩作用。胚胎形态发生过程中经常出现周期性的收缩脉冲。一种叫做细胞状态分裂器的模型涉及到交替细胞收缩和扩张,这是由一个在每个细胞顶端的双稳态细胞器发起的。有机器由微管和微丝组成,在机械的对立面。 它对由形态变化引起的局部机械扰动作出反应。然后,这些细胞会引发胚胎分化的波动,即收缩或扩张,超过了决定细胞类型的推定组织,然后是细胞分化。首先提出了细胞状态分裂器来解释轴突发育过程中的神经板形成,并将模型推广到所有形态发生过程。[12][13][14]
参考文献
- Thompson, D'Arcy Wentworth. . Cambridge University Press. 1917.
- Montell, Denise J, (PDF), Science, 5 December 2008, 322: 1502–1505 [2018-08-30], Bibcode:2008Sci...322.1502M, PMID 19056976, doi:10.1126/science.1164073, (原始内容 (PDF)存档于2014-11-28)
- Turing, A. M. . Philosophical Transactions of the Royal Society B. 1952, 237 (641): 37–72. Bibcode:1952RSPTB.237...37T. doi:10.1098/rstb.1952.0012.
- Hiscock, Tom W.; Megason, Sean G. . Cell Systems. 2015, 1 (6): 408–416. PMC 4707970. doi:10.1016/j.cels.2015.12.001.
- Kouros-Mehr, H.; Werb, Z. . Dev. Dyn. 2006, 235 (12): 3404–12 [2018-08-30]. PMC 2730892. PMID 17039550. doi:10.1002/dvdy.20978. (原始内容存档于2016-03-05).
- Montévil, Maël; Speroni, Lucia; Sonnenschein, Carlos; Soto, Ana M. . Progress in Biophysics and Molecular Biology. From the Century of the Genome to the Century of the Organism: New Theoretical Approaches. 2016, 122 (1): 58–69 [2018-08-30]. PMC 5563449. PMID 27544910. arXiv:1702.03337. doi:10.1016/j.pbiomolbio.2016.08.004. (原始内容存档于2018-10-09).
- Gilbert, Scott F. . 6th. Sunderland, Mass: Sinauer Associates. 2000 [2018-08-30]. ISBN 0-87893-243-7. (原始内容存档于2009-02-17).
- Hulpiau, P.; van Roy, F. . Int. J. Biochem. Cell Biol. February 2009, 41 (2): 349–69. PMID 18848899. doi:10.1016/j.biocel.2008.09.027.
- Angst, B.; Marcozzi, C.; Magee, A. . J Cell Sci. February 2001, 114 (Pt 4): 629–41. PMID 11171368.
- . Breast Cancer Res. 2004, 6 (1): 1–11. PMC 314442. PMID 14680479. doi:10.1186/bcr634.
- Sternlicht MD. . Breast Cancer Res. 2006, 8 (1): 201 [2018-08-30]. PMC 1413974. PMID 16524451. doi:10.1186/bcr1368. (原始内容存档于2014-10-06).
- Gordon R, Brodland GW. The cytoskeletal mechanics of brain morphogenesis. Cell state splitters cause primary neural induction. 页面存档备份,存于 Cell Biophys. 1987 Dec;11:177-238.
- Gordon, NK,Gordon,R. The organelle of differentiation in embryos: the cell state splitter 页面存档备份,存于 Theoretical Biology and Medical Modelling, 13(11) 2016
- Gordon NK, Gordon R Embryogenesis explained World Scientific Nov 2016, ISBN 978-981-4350-48-8
- (PDF). Annals of Clinical & Laboratory Science. 2008, 38 (4): 307–329 [2018-08-30]. PMID 18988924. (原始内容存档 (PDF)于2019-02-23).
延伸阅读
- Bard, J.B.L. . Cambridge, England: Cambridge University Press. 1990.剑桥,英国: 剑桥大学出版社。
- Slack, J.M.W. . Oxford: Wiley-Blackwell. 2013.牛津: 威利-布莱克威尔。
