脅空鳥龍屬
脅空鳥龍屬(学名:)又名拉鴻鳥,是種和鳥類非常相近的獸腳亞目恐龍,生存於晚白堊紀馬斯垂克階的馬達加斯加西北部,約7000萬年前到6500萬年前。脅空鳥龍的正模標本是一個部份骨骸(編號 UA 8656),發現於馬任加省的梅法拉諾組(Maevarano Formation),目前存放於安塔那那利佛大學(University of Antananarivo, UA)。脅空鳥龍是種小型恐龍,身長約70公分[3],第二腳趾有類似伶盜龍的鐮刀狀趾爪。
| 胁空鸟龙属 化石时期:白堊紀晚期, 70 Ma | |
|---|---|
 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龙总目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 兽脚亚目 Theropoda |
| 演化支: | 新兽脚类 Neotheropoda |
| 演化支: | 鸟吻类 Averostra |
| 演化支: | 坚尾龙类 Tetanurae |
| 演化支: | 俄里翁龙类 Orionides |
| 演化支: | 鸟兽脚类 Avetheropoda |
| 演化支: | 虚骨龙类 Coelurosauria |
| 演化支: | 暴盗龙类 Tyrannoraptora |
| 演化支: | 手盗龙形态类 Maniraptoromorpha |
| 演化支: | 手盗龙形类 Maniraptoriformes |
| 演化支: | 手盗龙类 Maniraptora |
| 演化支: | 廓羽盗龙类 Pennaraptora |
| 演化支: | 近鸟类 Paraves |
| 属: | †胁空鸟龙属 Rahonavis Forster et al, 1998b |
| 模式种 | |
| 奧氏脅空鳥龍 Rahonavis ostromi (Forster et al., 1998a) | |
| 異名[1][2] | |
| |
命名
脅空鳥龍的屬名中,rahona 在馬拉加斯語中意為「雲」或「威脅」,而 avis 在拉丁語中意為「鳥」,兩者意為「從空中威脅的鳥」;種名 ostromi 則是為了紀念已故美國古生物學家約翰·奧斯特倫姆(John Ostrom),他是恐龍文藝復興的主要推動者之一,對於恐龍和鳥類之間演化關係的研究有很深遠的貢獻。
脅空鳥龍的發現者最初將牠們命名為Rahona,但之後發現已有一種毒蛾科昆蟲名為Rahona,於是將牠們改名為Rahonavis[1][2]。
分類
脅空鳥龍的分類歷史有許多爭議,牠們可能屬於鳥綱演化支,或是相近的馳龍科。牠們尺骨的羽幹,使牠們最初被歸類於鳥類;然而,骨骸的其他部分則類似典型的馳龍科恐龍。原始鳥類與馳龍科的特性極為類似,而這群動物可能在演化過程中失去飛行能力,因此很難確定脅空鳥龍是否屬於鳥類。牠們的最初敘述者認為,脅空鳥龍可能是始祖鳥的近親,因此屬於鳥綱;脅空鳥龍的骨盆也顯示出飛行的適應演化,類似始祖鳥,但似乎是獨自發展出來的[4]。另一方面,彼得·馬克維奇(Peter Makovicky)與其同事則認為脅空鳥龍是南美洲馳龍科半鳥與鷲龍的近親,因此屬於半鳥亞科[5]。在2006年,諾瑞爾與其同事也认为脅空鳥龍屬於半鳥亞科,因此為半鳥的姐妹分類單元[6]。在2007年,愛倫·特納(Alan H. Turner)與其同事也认为脅空鳥龍屬於半鳥亞科,並為半鳥的近親[7]。
然而自2009年以来,许多传统上归类于驰龙科的类群被发现实际上更接近鸟类,如2013年研究[8]就指出胁空鸟龙比驰龙类更接近鸟类。
2014年勒菲弗等人的研究显示胁空鸟龙比始祖鸟更接近现代鸟类,是鸟翼类的成员[9]:
| 鳥翼類 Avialae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
發現與種
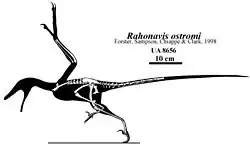

脅空鳥龍的化石是由紐約州立大學與塔那那利佛大學所組成的挖掘團隊,在1995年於貝立佛查村附近發現。這個地區的地層多覆蓋者密集的草,使得化石難以確認。當地的一片山坡在經過火災後,一個大型泰坦巨龍類的化石曝露出來。古生物學家在挖掘這個泰坦巨龍類化石時,也發現了脅空鳥龍的化石。脅空鳥龍的化石呈現天然狀態,由後肢、身體、部份尾巴、部分翅膀、以及肩膀所構成。脅空鳥龍的體型大於始祖鳥約1/5,相當於渡鴉的大小[10]。
由於缺乏相關近親的化石,因此難以確定脅空鳥龍的歸類,加拿大自然博物館的一節胸部脊椎骨,被發現於摩洛哥Kem Kem地區,年代為中白堊紀阿爾比階/森諾曼階,接近1億年前到9900萬年前,是最類似脅空鳥龍的化石,擁有較大的神經管,但卻乏脅空鳥龍具有的側腔,應該屬於不同的屬。雖然側腔的存在隨者不同物種、不同脊椎、以及不同個體而有所改變,但摩洛哥化石的時間與地點顯示牠們並不屬於脅空鳥龍[11]。
古生物學
有許多科學家的重建圖中,將脅空鳥龍重建為飛行的姿態,但目前仍無法確定牠們是否可以飛行。某些科學家認為脅空鳥龍的化石是個嵌合體,前肢骨頭來自於鳥類,其他部位骨頭來自於馳龍科,因此認為脅空鳥龍是個疑名[4]。在挖掘地點附近發現的Vorona,是個原始鳥類,顯示脅空鳥龍有可能是個嵌合體。而大部分科學家仍認為,前肢與其他部位的骨頭的發現位置,約於一張信封紙的大小,因此這些化石來自於單一動物,包含最初的研究人員在內。在2007年,路易斯·恰佩(Luis M. Chiappe)針對賴瑞·馬丁(Larry Martin)的嵌合體理論,指出賴瑞·馬丁誤將前肢與肩胛部位骨頭,敘述為具有更衍化的特徵[10]。路易斯·恰佩認為將脅空鳥龍與始祖鳥相比,脅空鳥龍的尺骨大而粗,尺骨上有明顯的羽莖瘤,顯示牠們具有比原始鳥類更大、更強壯的前肢,因此脅空鳥龍可以飛行。此外,脅空鳥龍的肩胛骨有韌帶附著的痕跡,顯示前肢可以獨自作出拍打的運動。恰佩認為脅空鳥龍可以飛行,但飛行能力較現代鳥類差[10]。
參考文献
- Forster, Catherine A.; Sampson, Scott D.; Chiappe, Luis M. & Krause, David W. (1998a): The Theropod Ancestry of Birds: New Evidence from the Late Cretaceous of Madagascar. Science 279(5358): 1915-1919. doi:10.1126/science.279.5358.1915 (HTML abstract)
- Forster, Catherine A.; Sampson, Scott D.; Chiappe, Luis M. & Krause, David W. (1998b): Genus Correction. Science 280(5361): 179.
- Holtz, Thomas R. Jr. (2008) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages Supplementary Information 页面存档备份,存于
- Geist, Nicholas R. & Feduccia, Alan (2000): Gravity-defying Behaviors: Identifying Models for Protoaves. American Zoologist 40: 664–675. PDF fulltext
- Makovicky, Peter J.; Apesteguía, Sebastián & Agnolín, Federico L. (2005): The earliest dromaeosaurid theropod from South America. Nature 437: 1007-1011. doi:10.1038/nature03996 (HTML abstract) Supplementary information
- Norell, M.A., Clark, J.M., Turner, A.H., Makovicky, P.J., Barsbold, R., and Rowe, T. (2006). "A new dromaeosaurid theropod from Ukhaa Tolgod (Omnogov, Mongolia)." American Museum Novitates, 3545: 1-51.
- Turner, Alan H.; Pol, Diego; Clarke, Julia A.; Erickson, Gregory M.; and Norell, Mark. (pdf). Science. 2007, 317: 1378–1381 [2007-09-21]. doi:10.1126/science.1144066. (原始内容存档 (PDF)于2010-11-05).
- Agnolín, F.L. and Novas, F.E. (2013). "Avian ancestors. A review of the phylogenetic relationships of the theropods Unenlagiidae, Microraptoria, Anchiornis and Scansoriopterygidae". SpringerBriefs in Earth System Sciences: 1–96.
- Lefèvre, U.; Hu, D.; Escuillié, F. O.; Dyke, G.; Godefroit, P. . Biological Journal of the Linnean Society. 2014, 113 (3): 790–804. doi:10.1111/bij.12343.
- Chiappe, L.M. . Sydney: UNSW Press.
- Riff, Douglas; Kellner, Alexander W. A.; Mader, Bryn & Russell, Dale (2002): On the occurrence of an avian vertebra in Cretaceous strata of Morocco, Africa. Anais da Academia Brasileira de Ciências 74(2): 367-368. doi:10.1590/S0001-37652002000200023 PDF fulltext
- Forster, Catherine A. & O'Conner (2000): The avifauna of the Upper Cretaceous Maevarano Formation, Madagascar. Society of Vertebrate Paleontology 20(3): 41A-42A.
- Schweitzer, Mary H.; Watt, John A.; Avci, Recep; Forster, Catherine A.; Krause, David W.; Knapp, Loren; Rogers, Raymond R.; Beech, Iwona & Marshall, Mark (1999): Keratin immunoreactivity in the Late Cretaceous bird Rahonavis ostromi. Society of Vertebrate Paleontology 19(4), 712-722. HTML abstract