馳龍科
馳龍科(学名:)又名奔龍科,是群類似鳥類的獸腳亞目恐龍。牠們是群體型中至小型的肉食性恐龍,繁盛於白堊紀。在非正式的用法裡,牠們通常被稱為「盜龍」(Raptor)[2],這個名稱因為電影《侏儸紀公園》(Jurassic Park)出現的伶盜龍而開始盛行。馳龍科的屬名在希臘文意為「奔跑的蜥蜴」,dromeus / 意為「奔跑」,而sauros / 意為「蜥蜴」。
| 驰龙科 化石时期:白堊紀[1], 143–66 Ma | |
|---|---|
 | |
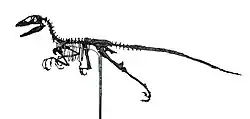
| 恐爪龍(後)與鷲龍(前)的骨骼 芝加哥菲爾德自然歷史博物館 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龙总目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 兽脚亚目 Theropoda |
| 演化支: | 新兽脚类 Neotheropoda |
| 演化支: | 鸟吻类 Averostra |
| 演化支: | 坚尾龙类 Tetanurae |
| 演化支: | 俄里翁龙类 Orionides |
| 演化支: | 鸟兽脚类 Avetheropoda |
| 演化支: | 虚骨龙类 Coelurosauria |
| 演化支: | 暴盗龙类 Tyrannoraptora |
| 演化支: | 手盗龙形态类 Maniraptoromorpha |
| 演化支: | 手盗龙形类 Maniraptoriformes |
| 演化支: | 手盗龙类 Maniraptora |
| 演化支: | 廓羽盗龙类 Pennaraptora |
| 演化支: | 近鸟类 Paraves |
| 演化支: | 真手盗龙类 Eumaniraptora |
| 下目: | †恐爪龙下目 Deinonychosauria |
| 科: | †驰龙科 Dromaeosauridae Matthew & Brown, 1922 |
| 模式种 | |
| †艾伯塔驰龙 Dromaeosaurus albertensis Matthew & Brown, 1922 | |
| 演化支 | |
| 異名 | |
| |
馳龍科的恐龍分布範圍可能極廣,化石在北美洲、歐洲、非洲、亞洲、南美洲以及南極洲等地[3]都有發現記錄。已知最原始的馳龍類恐龍記錄可能來自在英格蘭西南部被發現的牙齒化石,推測生活於侏羅紀巴通階[1],而後來演化並且持續存活到白堊紀末的馬斯垂克階,據此推算,馳龍類出現在地球上的時間超過1億年。
敘述
身體架構

馳龍科的獨特身體架構,有助於恐龍是種活躍且快速動物的理論,以及恐龍與鳥類是近親的理論。在羅伯特·巴克(Robert T. Bakker)替約翰·奧斯特倫姆(John Ostrom)的1969年專題論文所做的圖解中[4],將恐爪龍描繪成快速奔跑的姿態,這成為史上最具影響力的古生物學重建圖[5]。馳龍科具有大型頭部、鋸齒邊緣牙齒、狹窄口鼻部、眼睛向前,顯示具有一定程度的立體視覺[6]。如同大部分的獸腳類恐龍,馳龍科的頸部長,呈S狀彎曲,身體相當短。牠們的手臂長,某些物種的手臂可以在胸前交叉,手部相當大,上有三根長手指,第一指最短,第二指最長,手指上有大型指爪[7]。恥骨末端大,往尾巴基部延伸。第二腳趾上有大型、彎曲趾爪。尾巴修長,尾椎長而低平,第14節尾椎之後缺乏橫突(Transverse process)與神經棘[7]。
目前已發現部分馳龍科的身體覆蓋者羽毛,包含前肢與尾巴上的大型、片狀羽毛,可能所有的馳龍科都有羽毛。這個理論是在80年代中晚期首次出現,並在1999年得到證實。這些改變導致馳龍科在電影與重製模型中的形象不斷改變。
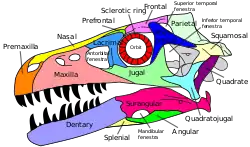
鑑定特徵
馳龍科具有以下特徵:額骨短而呈T字形,構成上顳孔的前緣、方骨的後突接觸方軛骨、背椎的椎體橫突對(Parapophyses)豎起、獨特的第二腳趾、人字形骨與尾椎的前關節突延長,延伸至前幾節脊椎、鳥喙骨上有關節盂孔[7]。
體型
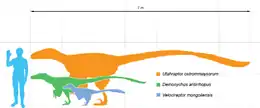
馳龍科是群小型到中型的恐龍,身長範圍從0.7公尺的大黑天神龍,到超過6公尺的猶他盜龍與阿基里斯龍[8][9]。楊百翰大學的某些猶他盜龍標本,身長可能達約11公尺。但這些化石還沒有經過詳細研究,經過詳細研究後可能會產生不同數值[10]。馳龍科可能兩次演化出大型體型,一次是馳龍亞科的猶他盜龍、阿基里斯龍,另一次是半鳥亞科的南方盜龍,其身長估計約5公尺。在英格蘭威特島發現的個別大型牙齒,可能代表某種大型馳龍科恐龍,體形接近於猶他盜龍。根據牙齒形狀,牠們可能屬於伶盜龍亞科。如果屬實,可能代表馳龍科分別三次演化出大型體型[11]。
目前所發現最原始的馳龍科是大黑天神龍,身長只有70公分,也是目前已知最小的馳龍科恐龍。大黑天神龍、小盜龍的發現,加上傷齒龍科的近鳥龍,顯示近鳥類的共同祖先可能非常的小,身長約65公分,體重約600到700公克;近鳥類包含馳龍科、傷龍科、以及鳥類[12]。
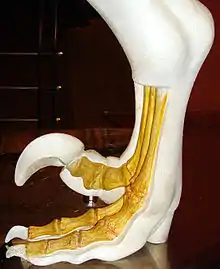
腳部
如同其他獸腳類恐龍,馳龍科以後肢行走。然而,大部分獸腳類恐龍以後肢的三根腳趾接觸地面;而馳龍科只有用第三、第四腳趾支撐身體重量,屬於雙趾型動物。從足跡化石顯示,牠們的第二趾明顯地後縮[13]。第二腳趾上有大型、鐮刀狀、彎曲趾爪,行走時會往後縮起,被認為是作為殺死獵物使用,或是攀爬樹木。真馳龍類的體型較大,第二趾爪的特化程度更為明顯[14]。真馳龍類的其中一屬,巴拉烏爾龍(Balaur)的第一趾、第二趾都朝向腳掌前方,這兩趾都向上後縮,都具有大型、鐮刀狀趾爪[15]。
尾巴
馳龍科具有長尾巴。大部分的尾椎有骨質、棒狀前關節突,某些物種的尾巴則具有骨化肌腱。在奧斯特倫姆的恐爪龍研究中,他提出這些特徵將尾巴硬挺,不易彎曲,惟有尾巴基部可靈活活動[4]。但是,蒙古伶盜龍的一個保存良好標本(編號IGM 100/986),尾巴向後彎曲呈S狀,關節沒有脫落,這顯示牠們的尾巴可做出一定程度的平行運動[16]。馳龍科的尾巴被假設在奔跑或滑翔時,作為穩定器或平衡器使用[16];在顧氏小盜龍與中國鳥龍的一個標本(編號 NGMC 91)的尾巴末端,有羽毛構成的小型、鑽石形扇狀物,可能作為氣動性上的穩定器,以及方向舵使用[17][18]。
羽毛
.jpg.webp)
有許多的證據顯示,馳龍科的身體覆蓋者羽毛。有些馳龍科化石的手部、手臂、尾巴,保存者長的正羽,而身體則保存者絨羽[19][20]。另外有些化石,雖然沒有保存羽毛,但前臂骨頭上則有突起處,可能在生前是羽毛的附著處[21]。整體而言,馳龍科的羽毛型態非常類似始祖鳥[20]。
第一種被發現有明顯羽毛證據的馳龍科恐龍是中國鳥龍,由徐星等人於1999年在中國發現[19]。許多其他的馳龍科化石曾發現羽毛覆蓋者身體,有些擁有完全發展的有羽毛翅膀。有些物種甚至顯示出後肢為第二對翼的證據,包括小盜龍與羽龍[20]。因為羽毛痕跡只會在細緻的沉積層中保存下來,有些發現於粗糙岩層中的化石保有了羽莖瘤,羽莖瘤是鳥類的飛羽附著點。脅空鳥龍與伶盜龍的化石都發現了羽莖瘤,顯示牠們的前肢具有羽毛。因此,大型、居住於地面的馳龍科恐龍極可能擁有羽毛,如同大部分的現代無法飛行鳥類仍保有牠們的飛羽[21][22]。雖然有些科學家主張,大型馳龍科失去了部份或全部的羽毛,但伶盜龍的羽莖瘤被視為是所有的馳龍科都具有羽毛的證據[23][24]。
與鳥類的關係
馳龍科屬於獸腳亞目,與早期的鳥類有許多共同特徵。有許多研究討論過馳龍科與鳥類的關係,隨者許多新證據的出現,馳龍科與鳥類關係的假設也不斷改變。在2001年,馬克·諾瑞爾(Mark Norell)與其同事研究大量的虛骨龍類化石,並提出馳龍科是鳥類的最近親,兩者與傷齒龍科共同屬於鳥翼類。諾瑞爾更指出,馳龍科是鳥翼類中的一個並系群[25]。在2002年,S.H. Hwang與其同事參考了諾瑞爾的研究與新發現化石,提出鳥類是馳龍科、傷齒龍科兩者的近親[8]。
古生物學界目前的共識多同意Hwang等人的研究,馳龍科與傷齒龍科是最近親,兩者共同組成恐爪龍下目。而恐爪龍下目是鳥類的姊妹分類單元,也是鳥類的最近親[26]。但古生物學界還沒有足夠證據可指出馳龍科(以及牠們的祖先)能否飛行或滑翔[27]。
相關理論
馳龍科傳統上被認為是非鳥類的恐龍,然而有些研究人員認為馳龍科也屬於鳥綱演化支。首先,馳龍科具有羽毛,符合傳統的鳥類定義-具有羽毛。羽龍意為「隱藏的飛行者」,牠們最初因其具有羽毛而被命名為鳥類,造成許多誤解[28]。

目前至少兩派研究人員提出馳龍科是從可飛行的祖先演化而來。其中一派主張鳥類先演化出現,喬治·奧利舍夫斯基(George Olshevsky)被認為是這理論的首位提出者[29]。葛瑞格利·保羅(Gregory S. Paul)則提出許多證據,指出馳龍科演化自會飛行的恐龍,可能類似始祖鳥。根據此理論,體型較大的馳龍科是在演化過程中失去飛行能力,如同紐西蘭的已滅絕恐鳥、以及鴕鳥[22]。在1988年,保羅提出馳龍科與現代鳥類的關係,比始祖鳥與現代鳥類的關係更為接近。但在2002年,保羅改提出馳龍科是始祖鳥的最近親[30]。
在2002年,S.H. Hwang與其同事提出小盜龍是最原始的馳龍科[8]。在2003年,徐星與其同事根據小盜龍的羽毛、翅膀等特徵,提出馳龍科的祖先能夠滑翔。而體型較大的馳龍科則在演化過程中失去滑翔能力,成為陸棲動物[20]。
在2002年,斯特芬·柯瑞克斯(Stephen Czerkas)敘述、命名羽龍。柯瑞克斯錯誤地判斷羽龍的羽毛只位在前肢與手部,將羽龍重建成雙翼恐龍;並認為羽龍較接近鳥類,而非滑翔動物。但是,日後有其他科學家提出羽龍可能是小盜龍的次異名。而柯瑞克斯也在他的著作中更正他的錯誤[31]。
另一派研究人員,例如賴利·馬丁(Larry Martin),則提出馳龍科與所有手盜龍類都不是恐龍。馬丁並提出鳥類與手盜龍類並非近親,但在2004年,馬丁修正他的理論,認為手盜龍類是鳥類的近親。但馬丁認為手盜龍類在演化過程中失去飛行能力,而鳥類演化自非恐龍的主龍類,而獸腳亞目的大部分成員並不屬於恐龍[32]。

在2005年,Mayr與Peters敘述了一個在懷俄明州瑟莫普利斯鎮發現的良好始祖鳥標本,發現始祖鳥的生理結構比原先認知的更類似恐龍。另外,他們還具有類似馳龍科的大幅彎曲第二趾爪、原始上頜、未反轉後趾。Mayr與Peters的種系發生學研究提出孔子鳥與小盜龍的親緣關係更為接近,而離始祖鳥較遠,使鳥翼類成為並系群演化支。他們更提出其近鳥類祖先能夠飛行或滑翔,而馳龍科與傷齒龍科在演化過程失去飛行或滑翔能力[34][35]。Corfe與Butler則以研究方法層面提出反對意見[36]。
在2007年,另有其他的理論出現。艾倫·特納(Alan Turner)與其同事命名了一種新馳龍科-大黑天神龍,大黑天神龍是最基礎、原始的馳龍科,比小盜龍更原始。大黑天神龍具有短手臂,無法滑翔。特納等人認為飛行能力演化自鳥翼類,而馳龍科的祖先無法飛行、滑翔。根據親緣分支分類法研究,原始的大黑天神龍顯示馳龍科的祖先無法飛行[37]。
古生物學

指爪與第二趾爪的功能
關於馳龍科第二腳趾的鐮刀狀趾爪的功能,目前存在着不同的意見。在1969年,當約翰·奧斯特倫姆(John Ostrom)敘述恐爪龍的鐮刀狀趾爪時,他將這些趾爪解釋為類似刀片的砍傷用武器,相當類似劍齒虎的犬齒,恐爪龍使用強壯的趾爪來挖開獵物的身體。在1987年,另有研究認為牠們會以第二趾爪獵殺大型角龍類恐龍[38]。這些解釋被應用到所有馳龍科恐龍身上。然而,菲利普·曼寧(Phillip Manning)等人認為這些趾爪並非當做鉤爪使用,並將它們重建為長橢圓形橫斷面,外圍包覆者角質層,而非原先認為的倒淚滴形橫斷面[39]。在曼寧的解釋中,第二趾爪可能在獵食大型獵食時,用來協助攀爬用,同時也是種刺傷武器。
在1970年,約翰·奧斯特倫姆曾參考鴕鳥與食火雞,發現這些鳥類會用較大型的第二趾爪攻擊獵物,進而產生恐爪龍會利用第二趾爪攻擊獵物的理論[4]。食火雞的第二趾爪可長達12.5公分[40]。奧斯特倫姆並引用一個1958年的鳥類論文,認為後肢與第二趾爪,可用來將獵物的身體撕開、將內臟挖出[41]。在近年,Chhristopher P. Kofron研究241件食火雞的攻擊紀錄,發現總計有1個人類、2隻狗的被攻擊死亡案例,但沒有證據顯示食火雞會用第二趾爪將獵物的內臟挖出[42][43]。相反地,食火雞將趾爪作為防衛武器,攻擊對牠們產生威脅的動物、或威嚇對方[40]叫鶴也具有較大的第二趾爪,卻是用在撕裂小型獵物,以適合吞嚥[44]。
在2009年,菲利普·曼寧等人著手研究第二趾爪與前肢指爪的功能。他們從生物力學方面開始研究,研究第二趾爪與前肢指爪在受力時,壓力與張力的分布與傳遞狀況。他們採用X光線成像技術,將一個伶盜龍的前肢指爪建立出3D立體圖像。接者,他們將雕鴞的指爪作為對照組。藉由比對發現,伶盜龍的前肢指爪相當適合用來攀爬。其尖銳的指爪尖端,適合穿刺、固定;而彎而寬的指爪基部,有助於平均地傳遞受力時的壓強[45]。
曼寧等人同時也研究第二趾爪的彎曲程度,並以現代鳥類與哺乳類作為對照組。根據早期的研究,趾爪的彎曲程度,與該動物的生活方式息息相關。趾爪大幅彎曲的動物,通常擅於攀爬;趾爪較筆直的動物,通常是在地面生存、活動的動物。曼寧等人發現,恐爪龍的第二趾爪,彎曲程度可達160度,屬於攀爬動物的範圍。同樣地,馳龍類的前肢指爪,彎曲程度也屬於攀爬動物的範圍[45]。
對於曼寧等人的研究,彼得·馬克維奇(Peter Mackovicky)有不同的看法。他認為小而原始的馳龍類(例如小盜龍)可能是攀爬動物,但晚期的大型馳龍類(例如阿基里斯龍)雖然具有大、彎曲的第二趾爪,但體型過大,已無法攀爬樹木。馬克維奇推測,晚期的大型馳龍類的第二趾爪,已失去攀爬功能,但會在獵食時,用來攻擊獵物[46]。
在2009年,菲力·森特(Phil Senter)發表一份馳龍科腳趾研究,提出部分物種可能會使用後肢趾爪挖開昆蟲的巢穴。某些小型馳龍科,例如:脅空鳥龍、鷲龍,可能有時會以昆蟲為食;而體型較大的馳龍科,例如:恐爪龍、內烏肯盜龍,也可能使用後肢趾爪挖開地表洞穴,以裡面的小型脊椎動物為食。但菲力·森特並沒有測試大幅彎曲的第二趾爪,在挖掘洞穴時是否有發生功能[47]。
在2011年的一項恐爪龍類趾爪研究,也認為第二趾爪無法作出有效的刺穿、劃開獵物身體的動作,並提出第二趾爪有固定獵物功能的理論,認為馳龍科可能以小型獵物為食。這些研究比對不同恐爪龍類、現代鳥類的腳掌結構、趾爪形狀。研究發現,許多掠食性現代鳥類也有大型第二趾爪,當牠們獵食小型獵物時,會使用腳掌、趾爪將獵物固定在地面上,用體重施壓於獵物,再用鳥喙吞食這些獵物,尤其是鷹科。不只是掠食性鳥類,其他掠食動物獵食時,也會用趾爪緊握、固定小型獵物[48]。研究發現恐爪龍的腳掌、腳趾應有大型肌腱,蹠骨短,第一趾(後趾)偏向後方,這些特徵很符合固定獵物功能的特徵。研究推測,當恐爪龍等進階型物種獵食時,可能會用第二趾爪攻擊、固定獵物到地面上,然後頭低下撕咬、吞食獵物,而尾巴具有平衡身體的功能[48]。牠們的牙齒後側才有大型、鉤狀邊緣,比較適合拔出獵物的肉體,而非撕裂。前肢強壯,有大型指爪,但覆蓋者長羽毛,當馳龍科獵食時,可能會將前肢張開,以伺機保持身體的平衡。以上特徵也支持固定獵物功能的理論[48]。
某些原始馳龍科、以及傷齒龍科的第二趾爪,還沒有演化成後期恐爪龍類的大型、鐮刀狀趾爪。牠們的腳趾關節允許作出更靈活的動作,而後期恐爪龍類的第二腳趾趾能作出向上後縮、向下的動作。這有可能是牠們以更小型動物為食,因此不用演化出大型趾爪、強壯腳掌以固定獵物[48]。研究人員推測,近鳥類可能共同擁有這些演化特徵,而現代鳥類則繼承這些特徵[49]。

群體獵食
曾經在腱龍的化石附近發現一個小群體的恐爪龍化石;腱龍是種大型草食性鳥腳下目恐龍。這發現被解釋成馳龍科恐龍以有秩序群體獵食的證據,類似某些現代哺乳類[50]。然而,並非所有古生物學家認為這證據具決定性,而後來的研究顯示恐爪龍較可能以無秩序的群體攻擊獵物。現代鳥類與鱷魚(馳龍科的近親們)顯示出很少協調的獵食行為;相反地,牠們通常是獨自獵食,或是被屍體所吸引,同一物種之內可能存在者這兩種不同的方式。以印尼的科摩多龍來說,牠們在群體進食的時候,最大型的個體會先進食,並攻擊企圖先進食的較小型個體;如果較小的個體死亡,將會遭到同類相食。當以這個現象來觀察原本被認為是群體獵食的馳龍科恐龍時,則呈現出同時符合科摩多龍與鱷魚的進食模式。這些地點所發現的恐爪龍骨骸都是來自於未成年體,而遺失的部份可能被其他恐爪龍所吞食,這不符合原先所認為的群體獵食行為[51]。
在2007年,科學家們在中國山東省發現了第一個大量的馳龍科足跡化石。這個足跡化石可能是由體型接近阿基里斯龍的大型馳龍科恐龍所留下,而且是由六個相近大小的個體,沿者海岸共同前進。這些個體相距約一公尺,朝同一方向緩慢前進。研究人員提出,這個足跡化石可證明馳龍科的某些物種過者群體生活。雖然該足跡化石並不能明確指出馳龍科的獵食行為,但無法排除牠們以群體獵食的可能性。此外,該發現還支持了馳龍科在行走時,第二腳趾後縮、離地的假設[13]。


飛行與滑翔能力
目前至少有兩種馳龍科恐龍被認為具有飛行或滑翔能力,脅空鳥龍、顧氏小盜龍。脅空鳥龍最初被歸類於鳥類,後來的研究將其歸類於馳龍科[52][53]。脅空鳥龍的長前肢被發現有羽莖瘤,羽莖瘤是飛羽的附著點,因此脅空鳥龍被認為可以動力飛行[54]。從結構來看,脅空鳥龍的前肢比始祖鳥更有力,並附有強壯的韌帶,可以作出拍動的動作,展開飛行。路易斯·齊亞比(Luis Chiappe)提出脅空鳥龍具有這些適應特徵,可以飛行,但飛行能力較現代鳥類差[55]。
顧氏小盜龍的前肢與後肢相當發達,科學家認為牠們可以採四翼方式滑翔。在2005年,薩克·查特吉(Sankar Chatterjee)提出顧氏小盜龍的四肢類似雙翼飛機,但是採前後排列方式。查特吉認為牠們在樹枝向下俯衝,以U字形的曲線滑翔,然後降落在地表或是其他的樹上;尾巴與後肢可用來控制方向與速度。此外,查特吉認為顧氏小盜龍有初步的飛行能力[17]。
分類系統
分類學
在1992年,W.D. Matthew與巴納姆·布郎(Barnum Brown)建立了馳龍亞科,當時只包含馳龍屬,屬於現在已廢止的恐爪龍科[62]。目前,馳龍科與傷齒龍科共同組成恐爪龍下目。
馳龍科的各亞科經常因為新的研究而改變內容,但大致上包含以下各演化支。而且許多屬的化石保存狀態不好,無法分類到任一亞科之中,例如最原始的大黑天神龍。
半鳥亞科(Unenlagiinae)是馳龍科中最原始的亞科[12]。半鳥亞科是個充滿謎題的演化支,是最少得到理論支持的亞科,其中有些或全部物種可能並不屬於馳龍科[63]。其中較大、生活於地面的物種,例如鷲龍與半鳥,顯示出強烈的飛行適應特徵,但牠們可能過大而不能飛行。另一個物種脅空鳥龍的體型非常小,擁有發展良好的翼,並有羽莖瘤(飛羽的附著點)的證據,牠們即有可能有飛行能力。
小盜龍類(Microraptoria)是馳龍科中第二原始的演化支。小盜龍類包含許多最小的馳龍科恐龍,並顯示出樹棲生活的證據。所有馳龍科的皮膚痕跡證據都來自於小盜龍類,所有小盜龍類都覆蓋者羽毛,並擁有發展良好的翼。如同半鳥亞科,有些物種可能擁有主動飛行能力。
伶盜龍亞科(Velociraptorinae)傳統上包含伶盜龍、恐爪龍,而新發現的白魔龍可能屬於伶盜龍亞科,而蜥鳥盜龍的分類位置仍未確定。
馳龍亞科(Dromaeosaurinae)通常包含體型中到大的物種,並擁有箱形的頭顱骨;其他的亞科通常擁有狹窄的口鼻部。
蜥鳥盜龍亞科(Saurornitholestinae)是尼克拉斯·朗里奇(Nicholas Longrich)與菲力·柯爾(Phil Currie)在2009年建立的新分類單元。西爪龍的發現,使得斑比盜龍不屬於小盜龍亞科。於是他們建立蜥鳥盜龍亞科,以包含蜥鳥盜龍與近親斑比盜龍[14]。





以下馳龍科各屬的分類表是根據賽里諾(2005年)、森特(2004年)、馬克維奇等人(2005)、諾瑞爾等人(2006年)、特納(2007年)等人、以及湯瑪斯·荷茲(2011年)的研究而來[26][37][52][53][64][65]。
- 馳龍科 Dromaeosauridae
- 欒川盜龍属 Luanchuanraptor
- 偵察龍属 Nuthetes
- 彭巴盜龍属 Pamparaptor
- 聯鳥龍属 Ornithodesmus
- 瓦爾盜龍属 Variraptor
- 火盜龍属 Pyroraptor
- 佛舞龍属 Shanag[14]
- 振元龍属 Zhenyuanlong
- 哈茲卡盜龍亞科 Halszkaraptorinae
- 哈茲卡盜龍属 Halszkaraptor
- 大黑天神龍属 Mahakala
- 胡山足龍属 Hulsanpes
- 半鳥亞科 Unenlagiinae[66]
- 小盜龍亞科 Microraptorinae[64]
( = 小盜龍類 Microraptoria ) - 真馳龍類 Eudromaeosauria[14]
- 恐爪龍属 Deinonychus
- 蜥鳥盜龍亞科 Saurornitholestinae[14]
- 伶盜龍亞科 Velociraptorinae[70]
- 冥河盜龍屬 Acheroraptor
- 依特米龍属 Itemirus[14]
- 臨河盜龍属 Linheraptor
- 伶盜龍属 Velociraptor
- 白魔龍属 Tsaagan
- 惡靈龍属 Adasaurus
- 馳龍亞科 Dromaeosaurinae[62]
- 阿基里斯龙属 Achillobator
- 驰龙属 Dromaeosaurus
- 达科他盗龙属 Dakotaraptor
- 似驰龙属 Dromaeosauroides
- 犹他盗龙属 Utahraptor
- 郊狼龙属 Yurgovuchia
系統發生學
在1998年,保羅·塞里諾(Paul Sereno)將馳龍科定義為一個演化支:包含馳龍,但不包含傷齒龍、似鳥龍、或家麻雀在內的最大天然族群演化支。馳龍科的不同亞科都曾被定義為演化支過,都常是以馳龍或其他次演化支作為定義基準點;舉例而言,馬克維奇將半鳥亞科定義為:在馳龍科中,所有親緣關係接近半鳥,而離伶盜龍較遠的所有物種。小盜龍類(Microraptoria)是唯一不是亞科的次演化支,森特與其同事在建立時,沒有加上亞科的字尾-inae,以避免被誤認,並獨立於馳龍科之外[26]。但塞里諾則提出不同的定義,並將小盜龍類改名為小盜龍亞科(Microraptorinae),歸類於馳龍科之內。但是,這個用法僅出現在他的TaxonSearch database網站,且未在正式文獻中出現過[64]。
以下演化樹是根據尼克拉斯·隆瑞奇與菲力·柯爾在2009年的研究,其中的亞科是採用賽里諾在2005年的定義[14]。
| 馳龍科 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 馳龍科 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 近鳥類 |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
備註:有些研究人員以及親緣分支分類法研究,尤其是針對馳龍科中的小型可飛行物種的研究,發現始祖鳥是馳龍科的早期成員。如果始祖鳥屬於馳龍科,根據國際動物命名委員會的規定,始祖鳥科將比馳龍科還具有優先權。因此若將始祖鳥包含在內,那馳龍科將改名為始祖鳥科。保羅·塞里諾是首批採用始祖鳥科,而不採用馳龍科的科學家之一[30]。
大眾文化

1993年的電影《侏儸紀公園》中所出現的伶盜龍,體型比該屬所發現最大的標本還大。实际上,这些“迅猛龙”是其近亲恐爪龙。 葛瑞格利·保羅(Gregory S. Paul)在他的書籍《Predatory Dinosaurs of the World》中,將恐爪龍列為伶盜龍的一種,並將該種命名為平衡伶盜龍(Velociraptor antirrhopus)[22],這個分類法後來遭到普遍地否決[7][73][74]。麥克·克萊頓將這个观点寫入他的《侏儸紀公園》系列小說當中,採用葛瑞格利·保羅的說法,并被应用到电影当中。在電影《侏儸紀公園》中出現的馳龍科恐龍,在當時是正確的描述,但現在看來則有許多方面是錯誤的,包含缺乏羽毛。但在《侏儸紀公園III》中則將這個錯誤糾正回來。
參考資料
- (英文)Metcalf, S.J., Vaughan, R.F., Benton, M.J., Cole, J., Simms, M.J. and Dartnall, D.L. . Proceedings of the Geologists’ Association. 1992, 103: 321–342.
- (英文)John Acorn. . University of Alberta Press. ´2007: 13. ISBN 0888644817.
- (英文)Case, J.A., Martin, J.E., and Reguero, M. (2007). "A dromaeosaur from the Maastrichtian of James Ross Island and the Late Cretaceous Antarctic dinosaur fauna." Pp. 1-4 in Cooper, A., Raymond, C., and Team, I.E. (eds.), Antarctica: a Keystone in a Changing World -- Online Proceedings for the Tenth International Symposium on Antarctic Earth Sciences, U.S. Geological Survey Open-File Report 2007-1047, SRP 083. U.S. Geological Survey, Washington, D.C.
- (英文)Ostrom, J. H. (PDF). Peabody Museum of Natural History Bulletin. 1969, 30: 1–165.
- (英文)Paul, Gregory S. (2006) "Drawing Dinosaurs"(2006). "Geotimes", January 2006. . [2007-12-18]. (原始内容存档于2008-05-02).
- (英文)Makovicky, Peter J.; Apesteguía, Sebastián; Agnolín, Federico L. . Nature. 2005, 437 (7061): 1007–1011. Bibcode:2005Natur.437.1007M. PMID 16222297. doi:10.1038/nature03996.
- (英文)Norell, M.A., and Makovicky, P.J. . Weishampel, D.B., Dodson, P., Osmólska, H. (编). 2nd edition. Berkeley: University of California Press. 2004: 196–210. ISBN 0-520-24209-2.
- (英文)Hwang, S.H.; Norell, M.A.; Ji, Q.; Gao, K. . American Museum Novitates. 2002, (3381): 44pp. hdl:2246/2870.
- (英文)Perle, A.; Norell, M.A.; Clark, J. . Contributions of the Mongolian-American Paleontological Project. 1999, 101: 1–105.
- (英文)Britt; Chure; Stadtman; Madsen; Scheetz; Burge. . Journal of Vertebrate Paleontology. 2001, 21 (3): 36A. doi:10.1080/02724634.2001.10010852.
- (英文)Naish, D. Hutt, and Martill, D.M. (2001). "Saurischian dinosaurs: theropods." in Martill, D.M. and Naish, D. (eds). Dinosaurs of the Isle of Wight. The Palaeontological Association, Field Guides to Fossils. 10, 242–309.
- (英文)Turner, A.S.; Hwang, S.H.; and Norell, M.A. (PDF). American Museum Novitates. 2007, 3557: 1–27 [2007-03-29]. doi:10.1206/0003-0082(2007)3557[1:ASDTFS]2.0.CO;2.
- (英文)Li, Rihui; Lockley, M.G., Makovicky, P.J., Matsukawa, M., Norell, M.A., Harris, J.D. and Liu, M. . Naturwissenschaften. 2007. (原始内容存档于2020-02-09).
- (英文)Longrich, N.R.; Currie, P.J. . PNAS. 2009, 106 (13): 5002–7. Bibcode:2009PNAS..106.5002L. PMC 2664043. PMID 19289829. doi:10.1073/pnas.0811664106.
- (英文)Z., Csiki; Vremir, M.; Brusatte, S. L.; and Norell, M. A. . Proceedings of the National Academy of Sciences of the United States of America. 2010, 107 (35): 15357–61. Bibcode:2010PNAS..10715357C. PMC 2932599. PMID 20805514. doi:10.1073/pnas.1006970107. Supporting Information
- (英文)Norell, Mark A.; & Makovicky, Peter J. . American Museum Novitates. 1999, 3282: 1–45.
- (英文)Chatterjee, S.; Templin, R.J. (PDF). Proceedings of the National Academy of Sciences. 2007, 104 (5): 1576–1580. Bibcode:2007PNAS..104.1576C. PMC 1780066. PMID 17242354. doi:10.1073/pnas.0609975104.
- (英文)Ji, Q., Norell, M. A., Gao, K-Q., Ji, S.-A., Ren, D. (2001). "The distribution of integumentary structures in a feathered dinosaur." Nature, 410(6832): 1084 - 1088.
- (英文)Xu, X., Wang, X.-L., and Wu, X.-C. . Nature. 1999, 401: 262–266. doi:10.1038/45769.
- (英文)Xing, X., Zhou, Z., Wang, X., Kuang, X., Zhang, F., and Du, X. (2003). "Four-winged dinosaurs from China." Nature, 421: 335–340.
- (英文)Turner, A.S.; Hwang, S.H.; and Norell, M.A. (PDF). American Museum Novitates. 2007, 3557: 1–27 [2007-03-29].
- (英文)Paul, Gregory S. (2002). Dinosaurs of the Air: The Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. 472 pp.
- (英文)Prum, R.; Brush, A.H. . The Quarterly Review of Biology. 2002, 77 (3): 261–295. PMID 12365352. doi:10.1086/341993.
- (英文)Turner, A.H.; Makovicky, P.J.; and Norell, M.A. (pdf). Science. 2007, 317 (5845): 1721.
- (英文)Norell, M. Clark, J.M., Makovicky, P.J. (2001). "Phylogenetic relationships among coelurosaurian theropods." New Perspectives on the Origin and Evolution of Birds: Proceedings of the International Symposium in Honor of John H. Ostrom, Yale Peabody Museum: 49-67.
- (英文)Senter, Phil; Barsbold, R.; Britt, Brooks B.; Burnham, David B. . Bulletin of the Gunma Museum of Natural History. 2004, 8: 1–20.
- (英文)Padian, Kevin; Dial, Kenneth P. . Nature. 2005, 438 (7066): E3; discussion E3–4. Bibcode:2005Natur.438E...3P. PMID 16292258. doi:10.1038/nature04354.
- (英文)Witmer, L.M. (2005) “The Debate on Avian Ancestry; Phylogeny, Function and Fossils.” In “Mesozoic Birds: Above the Heads of Dinosaurs”, pp. 3-30. ISBN 978-0-520-20094-4
- (英文)Olshevsky, George. (1994). "The birds first? A theory to fit the facts - evolution of reptiles into birds". Omni, June 1994. Volume 16 No. 9
- (英文)Paul, Gregory S. (1988). Predatory Dinosaurs of the World. New York: Simon and Schuster. 464 pp.
- (英文)Czerkas, S.A., Zhang, D., Li, J., and Li, Y. (2002). "Flying Dromaeosaurs", in Czerkas, S.J. (ed.): Feathered Dinosaurs and the Origin of Flight: The Dinosaur Museum Journal 1. Blanding: The Dinosaur Museum, 16-26.
- (英文)Martin, L.D. (PDF). Acta Geologica Sinica. 2004, 50 (6): 978–990.
- (英文)National Geographic News- Earliest Bird Had Feet Like Dinosaur, Fossil Shows - Nicholas Bakalar, December 1, 2005, Page 2. Retrieved 2006-10-18.
- (英文)Mayr, G; et al.. . Science. 2005, 310 (5753): 1483–1486. PMID 16322455. doi:10.1126/science.1120331.
- (英文)Mayr, G; Peters, SD. . Science. 2006, 313: 1238c. doi:10.1126/science.1130964.
- (英文)Corfe, IJ; Butler, RJ. . Science. 2006, 313 (5791): 1238b. PMID 16946054. doi:10.1126/science.1130800.
- (英文)Turner, Alan H.; Pol, Diego; Clarke, Julia A.; Erickson, Gregory M.; and Norell, Mark. (pdf). Science. 2007, 317: 1378–1381. doi:10.1126/science.1144066.
- (英文)Adams, Dawn (1987) "The bigger they are, the harder they fall: Implications of ischial curvature in ceratopsian dinosaurs" pg 1-6 in Currie, Philip J. and Koster, E. (eds) Fourth symposium on mesozoic terrestrial ecosystems. Tyrrell Museum, Drumheller, Canada
- (英文)Manning, P.L., Payne, D., Pennicott, J., Barrett, P.M., and Ennos, R.A. . Biology Letters. 2005, 2: 110–112. doi:10.1098/rsbl.2005.0395. (原始内容存档于2012-01-23).
- (英文)Davies, S. J. J. F. . New York: Oxford University Press. 2002. ISBN 0-19-854996-2.
- (英文)Gilliard, E. T. . Garden City, NY: Doubleday. 1958.
- (英文)Kofron, Christopher P. . Journal of Zoology. 1999, 249 (4): 375–381. doi:10.1111/j.1469-7998.1999.tb01206.x.
- (英文)Kofron, Christopher P. . Memoirs of the Queensland Museum. 2003, 49 (1): 335–338.
- (英文)Redford, Kent H.; Peters, Gustav. . Journal of Field Ornithology. 1986, 57 (4): 261–269. JSTOR 4513154.
- (英文)Manning, P.L.; Margetts, L.; Johnson, M.R.; Withers, P.J.; Sellers, W.I.; Falkingham, P.L.; Mummery, P.M.; Barrett, P.M.; Raymont, D.R.; 等. . The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology. 2009, 292 (9): 1397–1405. doi:10.1002/ar.20986.
- (英文). New Scientist. September 2009, (2725) [2009-09-15].
- (英文)Senter, P. . Bulletin of the Gunma Museum of Natural History. 2009, 13: 1–14.
- (英文)Fowler, D.W., Freedman, E.A., Scannella, J.B., Kambic, R.E. . PLoS ONE. 2011, 6 (12): e28964. doi:10.1371/journal.pone.0028964.
- Choi, Charles, , LiveScience, 14 December 2011
- (英文)Maxwell, W. D.; Ostrom, J.H. . Journal of Vertebrate Paleontology. 1995, 15 (4): 707–712.
- (英文)Roach, B. T.; D. L. Brinkman. . Bulletin of the Peabody Museum of Natural History. 2007, 48 (1): 103–138.
- (英文)Makovicky, Peter J., Apesteguía, Sebastián & Agnolín, Federico L. (2005). The earliest dromaeosaurid theropod from South America. Nature, 437: 1007–1011. doi:10.1038/nature03996
- (英文)Norell, M.A., Clark, J.M., Turner, A.H., Makovicky, P.J., Barsbold, R., and Rowe, T. (2006). "A new dromaeosaurid theropod from Ukhaa Tolgod (Omnogov, Mongolia)." American Museum Novitates, 3545: 1-51.
- (英文)Forster, Catherine A.; Sampson, Scott D.; Chiappe, Luis M. & Krause, David W. . Science. 1998a, (5358): pp. 1915–1919. doi:10.1126/science.279.5358.1915. (HTML abstract).
- (英文)Chiappe, L.M. . Sydney: UNSW Press.
- (英文)Feduccia, Alan, Lingham-Soliar, Theagarten, Hinchliffe, J. Richard. "Do feathered dinosaurs exist? Testing the hypothesis on neontological and paleontological evidence" "Journal of Morphology" 266:125-166
- (英文)Schmitz, L.; Motani, R. . Science. 2011, 332 (6030): 705–8. Bibcode:2011Sci...332..705S. PMID 21493820. doi:10.1126/science.1200043.
- (英文)Li, Quanguo, , Science, 9 March 2012, 335: 1215–1219, doi:10.1126/science.1213780
- (英文)Zelenitsky, D.K.; Therrien, F.; Kobayashi, Y. . Proceedings of the Royal Society B. 2009, 276 (1657): 667–673. PMC 2660930. PMID 18957367. doi:10.1098/rspb.2008.1075.
- (英文)Zelenitsky, D.K.; Therrien, F.; Ridgely, R.C.; McGee, A.R.; Witmer, L.M. . Proceedings of the Royal Society B. 2011,. in press (1725): 3625. doi:10.1098/rspb.2011.0238.
- (英文)Rothschild, B., Tanke, D. H., and Ford, T. L., 2001, Theropod stress fractures and tendon avulsions as a clue to activity: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 331-336.
- (英文)Matthew, W. D.; Brown, B. . Bulletin of the American Museum of Natural History. 1922, 46: 367–385. hdl:2246/1300.
- (葡萄牙文)Agnolin, F.L.; Novas, F.E. (PDF). Anais da Academia Brasileira de Ciências. 2011, 83: 117–162 [2011-04-23]. doi:10.1590/S0001-37652011000100008.
- (英文)Sereno, P. C. . Systematic Biology. 2005, 51: 1–25.
- Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix.
- Bonaparte, (1999).
- (英文)Novas, F.E., Pol, D., Canale, J.I., Porfiri, J.D., and Calvo, J.O. (2008). "A bizarre Cretaceous theropod dinosaur from Patagonia and the evolution of Gondwanan dromaeosaurids." Proceedings of the Royal Society B, doi:10.1098/rspb.2008.1554
- (英文)Novas and Agnolin, (2004). "Unquillosaurus ceibalii Powell, a giant maniraptoran (Dinosauria, Theropoda) from the Late Cretaceous of Argentina." Rev. Mus. Argentino Cienc. Nat., n.s. 6(1): 61-66.
- (英文)Zheng, X., Xu, X., You, H., Zhao, Q. and Dong, Z. (2009). "A short-armed dromaeosaurid from the Jehol Group of China with implications for early dromaeosaurid evolution." Proceedings of the Royal Society B, published online before print August 19, 2009. doi:10.1098/rspb.2009.1178
- (俄文)Barsbold, R. (1983). "O ptich'ikh chertakh v stroyenii khishchnykh dinozavrov. ["Avian" features in the morphology of predatory dinosaurs]." Transactions of the Joint Soviet Mongolian Paleontological Expedition 24: 96-103. [原文為俄文] Translated by W. Robert Welsh, copy provided by Kenneth Carpenter and converted by Matthew Carrano. PDF fulltext WebCite的存檔,存档日期2007-07-20
- (英文)Senter P., Kirkland J.I., DeBlieux D.D., Madsen S., Toth N. . PLoS ONE. 2012, 7 (5): e36790. doi:10.1371/journal.pone.0036790.
- (英文)Turner, A. H.; Makovicky, P. J.; Norell, M. A. (2012). "A Review of Dromaeosaurid Systematics and Paravian Phylogeny". Bulletin of the American Museum of Natural History 371: 1. doi:10.1206/748.1
- (西班牙文)Pérez-Moreno, B.P.; J. L. Sanz, J. Sudre and B. Sigé. . Dinosaurs and Other Fossil Reptiles of Europe, Second Georges Cuvier Symposium, Montbéliard; Revue de Paléobiologie, Volume spécial. 1994, 7: 173–188.
- (英文)Currie, P. J. . Journal of Vertebrate Paleontology. 1995, 15 (3): 576–591. (abstract 的存檔,存档日期2007-09-27.)
外部連結
| 维基物种中的分类信息:馳龍科 |
| 维基共享资源中相关的多媒体资源:馳龍科 |
- (英文)馳龍科的列表、地理與時代分布 - DinoData
- (英文)馳龍科的簡介 - 加州大學柏克萊分校的古生物博物館網站
- (英文)[馳龍科的簡略演化樹,以及各亞科、各屬的概述] - Thescelosaurus
- (英文)馳龍科的定義歷史 - TaxonSearch
