合弓綱
合弓綱(Synapsida)意為「固定的顴弓」,也被稱成獸形綱(Theropsida),是羊膜動物的一綱,包含羊膜動物中所有與哺乳類關係較近的物種[1]。合弓綱是羊膜動物的兩個主要演化支之一,另一個演化支為蜥形綱(Sauropsida)。
| 合弓綱 化石时期:312–80 Ma 石炭紀晚期到白堊紀晚期 (不包含哺乳類的狀況) | |
|---|---|
 | |
| 合弓綱的代表种类: 杯鼻龙(属于卡色龙科)、异齿龙(属于楔齿龙科)、狼蜥兽(属于丽齿兽亚目)、麝足兽(属于恐头兽亚目貘头兽科)、獭形狸尾兽和Adelobasileus(属于犬齿兽亚目哺乳形类)、虎和澳洲针鼹(属于哺乳形类的一支后代:哺乳类)。 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 总纲: | 四足总纲 Tetrapoda |
| 演化支: | 爬行形类 Reptiliomorpha |
| 演化支: | 羊膜动物 Amniota |
| 纲: | †合弓纲 Synapsida Osborn, 1903 |
| 目 | |
| 異名 | |
| |
在傳統的分類法中,合弓綱曾是爬行動物的一個亞綱,因合弓綱動物在头骨的兩側各有一個下颞孔,故又名單孔亞綱、下孔亞綱,或名似哺乳爬行動物[2][3]。在此定義下,哺乳動物起源於爬行動物,但由於分類定義的變化,這種觀點目前是不對的。
合弓綱出現在3億800萬年前的晚石炭紀,並成為二叠纪早到晚期的優勢陸地動物,數量眾多且多樣化。二疊紀-三疊紀滅絕事件發生時,許多早期合弓類動物消失,少數物種存活到三叠纪,而主龍類爬行動物迅速成為三疊紀的優勢陸地動物。合弓綱在三叠纪以后渐趋绝灭,只有少數物種存活到白堊紀。現今的哺乳類是合弓綱的後代。
不斷改變的分類
在20世紀的早期,合弓類動物被分為爬行綱中的四個亞綱之一,這是根據顳顬孔的形態、位置所定的分類。這些位於頭部兩側的下顳孔,可使下頜肌肉附著在上面,造成更有力的咬合。在傳統的觀念裡,單孔亞綱是逐漸演化成哺乳動物的一支爬行動物,因為牠們逐漸演化出哺乳類的特徵,牠們也稱為似哺乳爬行動物[2][3]。
這傳統的分類持續用到80年代晚期。在90年代被親緣分支分類法取代,這是根據共同祖先與共同祖先之所有後代的分法,只有單系群才被承認為有效的分類單元或演化支。因為合弓類演化出哺乳動物,哺乳動物被分在合弓類演化支之內。但在一般狀態下,仍將合弓綱用來包含此演化支中的非哺乳綱物種。而爬行綱被蜥形綱取代,只包含無孔亞綱、雙孔亞綱;調孔亞綱被認為是複系群,不再被使用。因此,似哺乳爬行動物也較少被使用。
合弓類(不包含哺乳類)通常依照演化的程度,被分為兩個分類:盤龍目、獸孔目。盤龍目至少包含六科,因為演化程度較為原始而被歸類在一起,因此是個非天然的並系群[3]。盤龍目可分為卡色龍亞目、真盤龍亞目,真盤龍亞目演化出獸孔目、哺乳綱,也是一個並系群。
特徵
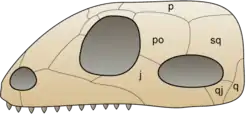
在合弓類的頭顱兩側,眼窩後面各演化出一個顳顬孔。顳顬孔可能用來增加下頜肌肉的附著處。雙孔亞綱爬行動物則演化出兩個顳顬孔。
合弓類是第一批演化出異型齒牙齒的四足動物。這些不同形態牙齒包括:犬齒、臼齒、以及門齒。某些离片椎类與早期無孔類爬行動物,上頜的第一對牙齒較大,已經出現牙齒異化的演化趨勢。蜥形綱演化支在演化過程中失去這個特徵,合弓類的牙齒則繼續特化發展。早期合弓動物的上頜有二或三對大型犬齒,獸孔類的上頜則有一對大型犬齒。下頜的犬齒則是在哺乳類時期演化出現。
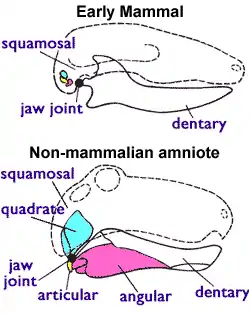
大部分古動物學家根據下頜特徵,來分辨合弓動物與爬行動物。許多科學家也根據下頜的特徵變化,來分辨合弓類演化至哺乳類的各個階段。合弓動物、史前與現代爬行動物的下頜是由多塊小骨頭構成,包含:齒骨、關節骨、與其他骨頭;而現代哺乳動物的下頜是由一塊齒骨構成。早期合弓動物有多塊下頜骨頭。隨者牠們的演化,這些下頜骨頭縮小,並且逐漸移入耳內,形成中耳的骨頭。合弓動物的耳部骨頭僅有鐙骨,哺乳動物則發展出鎚骨、砧骨、鐙骨。鎚骨是由關節骨演化而成,而砧骨是由方骨演化而成[4]。
哺乳動物的頜部關節是由齒骨、鱗狀骨相接而成,齒骨連結至鱗狀骨上的關節盂(Glenoid cavity)。其他的有頜類脊椎動物(包含爬行動物與合弓類)的頜部關節,是由下頜的關節骨與頭部的方骨相接而成。在合弓類演化成哺乳類的過程中,下頜的骨頭漸少,最後只剩下齒骨;下頜不再連接至鱗狀骨,改連接至方骨、以及往後退的關節骨。
在合弓類演化成哺乳類的過程中,合弓類演化出次生顎,分隔口腔與鼻腔。早期合弓類的上頜骨兩側已具有初步的次生顎,形狀成U形,口腔與鼻腔仍然連接。晚期合弓類的次生顎逐漸成C形,將口腔、鼻腔分隔,`形成腭骨。上頜骨也完全的癒合。在一個早期的真獸孔類化石中,已見初步的次生顎。後期的三尖叉齒獸,已具有完全閉合的次生顎[5]。
長期以來,合弓類的外表組織構造為何,仍處在爭議中。原始爬行動物的外皮可能相當薄,缺乏真皮層[6]。現存爬行動物的外表由角質鱗片、鱗甲所構成,而現存哺乳動物的外表則是表皮、真皮、皮下組織所構成的皮膚,只有少數哺乳動物仍保有鱗片。如同哺乳類,合弓類動物的表皮缺乏鱗片,具有皮腺。
目前仍不清楚合弓類演化至哺乳類的過程中,外皮是如何演化改變。但根據化石顯示,盤龍目的腹部具有鱗片,如同原始四足類。盤龍目的鱗片是由真皮衍化而成的鱗片,類似現代鱷魚、烏龜;而蜥蜴、蛇的的鱗片是由表皮形成的[7]。盤龍目的身體上半側則可能沒有鱗片。在外表上,盤龍目看起來像是沒有鱗片的爬行動物,沒有毛髮與鱗片。古動物學家多藉由骨骼特徵,來分辨盤龍類、其他原始爬行動物。

由於化石很少保存軟組織,目前還不清楚哺乳動物何時演化出毛髮與乳腺。在沒有發現皮膚痕跡的化石,必須藉由骨骼特徵,來判斷史前合弓動物在哪個時期需要毛髮隔絕熱量、具有乳腺。進階型獸孔類的外表可能由毛髮、鱗片、乳腺所構成,類似某些現代原始哺乳動物。
有些合弓動物(包括哺乳類)可能是溫血動物;但早期合弓類(例如盤龍目)可能大部分是冷血動物。進階型犬齒獸類具有次生顎、直立的四肢步態、以及毛髮等特徵,顯示牠們已經相當類似哺乳類,而且已有高代謝率。某些獸孔類化石的皮膚痕跡,也證實毛髮的存在[8]。
演化史

二疊紀
盤龍目是第一批成功的羊膜動物,散佈各地並多樣化發展,直到牠們成為石炭紀晚期到二疊紀早期的陸地優勢動物。牠們通常被分為兩支:真盤龍亞目、卡色龍亞目。牠們的體積龐大、四肢成躺臥狀態、腦部小,可能是群變溫動物。牠們是那個時代陸地上最大的動物,某些物種可達3公尺長。有些後期盤龍類,擁有大型帆狀物,可以幫助提升他們體溫,例如異齒龍、基龍。大部分盤龍類在二疊紀中期之前消失,少數殘存的物種存活到二疊紀中期。
獸孔目是一群更為進步的合弓類動物,出現在二疊紀空谷阶,成為二疊紀後半期的陸地優勢動物,並把合弓纲的多样性推上了一个高峰[10]。獸孔目一共成為優勢動物兩次:一次在二疊紀,一次在新生代(哺乳類)。在二疊紀中期到晚期,獸孔目是最具多樣性與數量最多的陸地動物,包括植食性、肉食性動物,體型從小如老鼠(例:羅伯特獸),體型大的可以以公噸計算(例:麝足獸)。在繁盛了数千萬年之後,這些成功的動物幾乎因為2億5100萬年前的二疊紀-三疊紀滅絕事件而滅絕了,這個地球歷史上最大的生物滅絕事件,可能跟西伯利亞暗色岩火山爆發事件有關。
三疊紀

盤龍目早已在二疊紀中消亡,在二疊紀-三疊紀滅絕事件之後,只有少數獸孔目存活下來,並且成功在三疊紀早期活着,包括:水龍獸、犬頜獸,後者出現在三疊紀中期,牠們與早期主龍形類(原先被命名為槽齒目,已经不在現代分類法中使用)同時存在。有些早期主龍形類是小型而敏捷的動物,例如派克鱷;大型主龍形類可以比肩大型獸孔目,例如引鱷。
三疊紀的獸孔目包括三個演化支。喙狀嘴的植食性二齒獸類(例如:水龍獸、肯氏獸),有些物種可身長到以公噸計算。越來越類似哺乳類的犬齒獸類,分植食性、肉食性、食蟲性動物,包括出現在奧倫尼克階的真犬齒獸類,早期的代表是犬頜獸。第三個是獸頭類,僅存活到三疊紀中期。

在三疊紀晚期,犬齒獸類的體型逐漸的變小,並且更類似哺乳類,跟體積逐漸变大的二齒獸類不同。最小、最進化的犬齒獸類只有鼩鼱般大小,生存在三疊紀晚期,大約2億年前,成為哺乳類的共同祖先。
因為氣候變遷、植被改變、生態位競爭、或者綜合以上因素,大部分的大型犬齒獸類在卡尼階時代消失,后来的三疊紀-侏儸紀滅絕事件不仅消滅了所有大型非恐龍主龍類,还消滅了二齒獸類。二齒獸類的生態位置在剩下的中生代時期被部分恐龍取代。存活下來的合弓纲很小,體型從鼩鼱到類似獾的強壯爬獸(Repenomamus)。
侏儸紀之后
隨著早期獸孔目-犬齒獸類-真犬齒獸類-哺乳類的階段性進化,主要的下頜骨頭齒骨,逐漸取代其他下頜的骨頭。所以,下頜逐漸形成只有一個大型骨頭,幾塊小型的下頜骨頭逐漸移動到內耳裡,使得聽覺更為靈敏。
在侏儸紀與白堊紀期間,這些存活的非哺乳犬齒獸類非常小,例如:三瘤齒獸。沒有任何犬齒獸類長得比貓還大。大多數侏儸紀與白堊紀的犬齒獸類是草食性動物,有些是肉食性的。三稜齒獸科在侏儸紀末第一次出現。牠們是肉食性動物,並且存活到侏儸紀中期。三瘤齒獸科跟三稜齒獸科同時出現,但為草食性動物,三瘤齒獸科在早白堊紀末期滅絕。二齒獸類被認為在三疊紀末期滅絕,但還是有證據顯示這演化支持續存活者。在岡瓦那大陸的白堊紀地層中發現新的二齒獸類化石。這是個化石記錄中忽隱忽現的物種的例子(俗稱的拉撒路物種)。
現在,有大約5,400個種的合弓纲存活者,牠們現名為哺乳動物,包括水生(鯨魚)、飛行(蝙蝠)動物,還有到目前為止已知最大的動物(藍鯨),以及人類。除了單孔目以外,大部分的哺乳動物已演化成胎生動物,直接產下幼體,而非產卵。
合弓動物進化成哺乳類的原因,可能是因為逐漸繁盛的恐龍在夜晚並不佔優勢,因此合弓類必須在晚上行動。為了在夜間生存,原始的哺乳類必須在夜間增加牠们的代謝率以維持體溫,這意味者更快地消耗食物(一般被認為是昆蟲)。為了促進快速消化,原始的哺乳類演化出咀嚼與特化的牙齒來協助咀嚼。四肢也演化到身體底下,而不是身體兩側[11]。這讓原始哺乳類能更快改變方向以抓住小型獵物。與快速奔跑的獵食者相比,原始哺乳類改採機動性的策略[8]。
分類
參考資料
- Laurin, Michel, and Robert R. Reisz (2007). Synapsida: Mammals and their extinct relatives 页面存档备份,存于. Version 06 April 2007. The Tree of Life Web Project.
- Benton, Michael J. 3rd ed. Oxford: Blackwell Science Ltd. 2004. ISBN 0632056371.
- Carroll, R. L. . New York: WH Freeman & Co. 1988. ISBN 0716718227.
- Salentijn, L. Biology of Mineralized Tissues: Prenatal Skull Development, Columbia University College of Dental Medicine post-graduate dental lecture series, 2007
- Hopson, James A. . The American Biology Teacher. 1987, 49 (1): 16–26.
- Hildebran, M. & Goslow, G. (2001): Analysis of Vertebrate Structure. 5th edition. John Wiley & sons inc, New York. 635 pages ISBN 978-0-471-29505-1
- Carroll, R.L. (1969). "Problems of the origin of reptiles." Biological Reviews, 44: 393-432.
- Hoyt, Donald F. (1997). Synapsid Reptiles 页面存档备份,存于. Zoo 138 Vertebrate Zoology Home Page.
- Lambert, David (2001). Dinosaur Encyclopedia. ISBN 978-0-7894-7935-8.
- Brocklehurst, Neil; Kammerer, Christian F.; Fröbisch, Jörg. . Paleobiology. 2013, 39 (3): 470–490. ISSN 0094-8373. doi:10.1666/12049 (英语).
- Bramble, D. M.; Jenkins, F. A. . Science. 1993, 262 (5131): 235–240. doi:10.1126/science.8211141.
- Colbert, E. H. 2nd ed. New York: John Wiley & Sons Inc. 1969. ISBN 0471164666.
相關條目
外部連結
- 合弓動物 - 盤龍目 - at Palaeos
- Transitional Vertebrate Fossils页面存档备份,存于 - includes description of important transitional genera from reptile to mammal
| 维基物种中的分类信息:合弓綱 |
