魚鰓
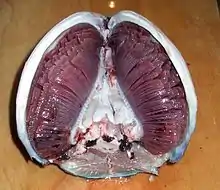
魚鰓是魚類的鰓,位於其咽部左右。鰓中有蛋白絲結構,其中含有毛細血管以交換氧氣與二氧化碳。它們通過將含有氧氣的水由嘴吸入鰓中來獲取之,一些魚類毛細血管中血液流向與水流相反,可以造成逆流交換,最終將含氧量減少的水從鰓中排出。
.jpg.webp)
一些魚類,諸如鯊魚和七鰓鰻,擁有不止一對鰓,這與硬骨魚大相徑庭。幼年多鰭魚也有多個鰓,是一種同兩栖類幼體相似的原始結構。
呼吸

首先並不是所有的魚都要用鰓呼吸,不過可以直接呼吸空氣的魚也分兩種,例如非洲肺魚是必須要隔一段時間就需要呼吸空氣的,否則它們將窒息而死。而像下口鯰這一類的魚虽然可以直接呼吸空氣,但这对它们来说并非是必要的。大部份可呼吸空氣的魚都屬於後一種,因為這樣可以最大程度地避開水面上的捕食者。[1] 一些鳗鲡科魚類還可以直接通過皮膚吸收氧氣。[1]
除此之外,基本上所有的有脊椎魚類都是要用鰓呼吸的,其每一個鰓頁都有鰓弓支撐。[2] 典型的魚鰓位於咽部兩側,通過鰓裂與外部溝通。鰓中的蛋白絲結構稱為鰓小片,可以增大魚鰓與水中氧氣的接觸面積。[3] 當一條魚呼吸時,它會規律地將水從嘴中吸入,并強迫其通過鰓流出。硬骨魚有三對鰓弓,軟骨魚則有五到七對,原始的無腭魚也有七對。而有脊椎魚類的遠祖們則更多,一些脊索動物門動物甚至有五十對鰓。[4]
蛋白絲組成的大受氧面積對魚類換氣至關重要,因為水中溶解的氧氣只有很少一部份且速度極慢,標準狀況下水中每立方米僅僅含有二百五十克氧氣,一升淡水中只有八立方厘米氧氣,而同樣條件下空氣中則含有二百一十立方厘米,[5] 於此同時水的粘性是空氣的一百多倍,密度則是七百多倍。[5] 不過隨著逐漸適應,大多數魚類已不能離水存活,水的密度可以保證其鰓不相互粘連而失去呼吸能力。[5]
高等脊椎動物不會發展出鰓,雖然它們會在生前發育階段長出鰓弓,但最終都會變成其他器官諸如甲狀腺、喉部、聽小骨[4]的基礎。
硬骨魚
硬骨魚的鰓位於鰓蓋下,鰓蓋可以調整咽部水壓以保證有合適的水量通過腮部,因此並不需要“衝撞換氣”(即不停地游動),其嘴內部的閥門構造也可以防止水倒流而出。[4]
硬骨魚的鰓弓之間沒有隔膜,每個鰓弓支撐單獨的鰓條,一些種類的魚還有鰓耙。幾乎大部份的原始硬骨魚都缺少噴水孔,而保有與之相連的假鰓,位於鰓蓋基部,大部份又已然退化,沒有保留一絲的鰓狀構造。[4]
海生硬骨魚也用鰓來排出電解質,同時它們的鰓可能還與其體內液的同渗容摩有關。鹹水比體內液稠一些,因此鹹水魚在呼吸時會失去一部份體內的水份。爲了重新獲取,它們會喝下大量海水並排除鹽分。而淡水因為比體內液濃度低,淡水魚在呼吸時反而可以吸收到水份。[4]
一些原始硬骨魚同兩栖動物一樣,幼體擁有從鰓弓上分出的外鰓。[6] 成年之後退化,硬骨魚的外鰓由鰓本身控制,兩栖類則由肺部控制。在硬骨魚向四足動物進化的早期階段,內部鰓系統就已經完全消失了。[7]
軟骨魚


鯊魚和新鰩目通常有五個鰓裂,直接與體外聯通,一些更原始的鯊魚則有六到七對。相鄰的鰓裂由軟骨鰓弓上長出的頁裝隔膜分割開來。單獨的鰓薄片位於隔膜兩側,鰓弓基部可能也支撐著鳃耙,用以過濾水。[4]
噴水孔位於第一對鰓裂的後部,它們還有假鰓。[4] 噴水孔可能與高等脊椎動物的耳孔是同源的。[8]
大部份鯊魚依靠“衝撞換氣”,即通過不斷迅速遊動強迫水從腮部流過。一些底棲或者游速緩慢的軟骨魚,例如新鰩目的一些魚類,有更大的噴水孔,它們通過噴水孔來吸入水,而不是靠嘴。[4]
七鰓鰻和八目鰻
七鰓鰻和八目鰻沒有鰓裂,它們的鰓部有一個球形囊,並有圓孔與外界聯通,每個球性囊中有兩個鰓。一些種類的圓孔已經融合,形成一個鰓蓋。七鰓鰻有七对這樣的囊,而不同種的八目鰻可能由六到十四對。八目鰻的球形囊在內部與咽相接。[4]
寄生蟲
魚鰓是許多皮外寄生蟲(寄生於鰓的外部)的寄生地,最常見的是單殖亞綱生物和一些種類的橈足動物,其數量龐大。[9]其他寄生蟲還包括蛭、颚水虱类幼蟲。[10]體內寄生蟲(寄生於鰓的內部)有被囊的囊双科吸蟲成蟲。[11]Huffmanela屬的少數線蟲,諸如Huffmanela ossicola會寄生在魚鰓骨中,[12]涡虫纲的被囊寄生蟲Paravortex也是其中一種。[13]寄生蟲有時是致命的,密集的寄生在鰓部可能會造成魚隻缺氧而死。
參見
- 水下呼吸
- 書肺
- 鳃耙
- 腮裂
- 人工腮
參考文獻
- (PDF). Copeia. 1998, (3): 663–675 [25 June 2009].
- Scott, Thomas. . Walter de Gruyter. 1996: 542. ISBN 978-3-11-010661-9.
- Andrews, Chris; Adrian Exell, Neville Carrington. . Firefly Books. 2003.
- Romer, Alfred Sherwood; Parsons, Thomas S. . Philadelphia, PA: Holt-Saunders International. 1977: 316–327. ISBN 0-03-910284-X.
- M. b. v. Roberts, Michael Reiss, Grace Monger. . London, UK: Nelson. 2000: 164–165.
- The American Naturalist (Essex Institute). 1957, 91: 287. 缺少或
|title=为空 (帮助) - Clack, J. A. (2002): Gaining ground: the origin and evolution of tetrapods. Indiana University Press, Bloomington, Indiana. 369 pp
- Laurin M. (1998): The importance of global parsimony and historical bias in understanding tetrapod evolution. Part I-systematics, middle ear evolution, and jaw suspension. Annales des Sciences Naturelles, Zoologie, Paris, 13e Série 19: pp 1-42.
- Kearn, G. C. (2004). Leeches, Lice and Lampreys. A natural history of skin and gill parasites of fishes. Dordrecht: Springer.
- Grutter, A. S. 1994: Spatial and temporal variations of the ectoparasites of seven reef fish species from Lizard Island and Heron Island, Australia. Marine Ecology Progress Series, 115, 21-30.
- Pozdnyakov, S. E. & Gibson, D. I. (2008). Family Didymozoidae Monticelli, 1888. In R. A. Bray, D. I. Gibson & A. Jones (Eds.), Keys to the Trematoda, Vol. 3 (pp. 631-734). London: CAB International and The Natural History Museum.
- Justine, J.-L. 2004: Three new species of Huffmanela Moravec, 1987 (Nematoda: Trichosomoididae) from the gills of marine fish off New Caledonia. Systematic Parasitology, 59, 29-37.
- Cannon, L. R. G. & Lester, R. J. G. 1988: Two turbellarians parasitic in fish. Diseases of Aquatic Organisms, 5, 15-22.
外部連結
- Fish Dissection - Gills exposed Australian Museum. Updated: 11 June 2010. Retrieved 16 January 2012.
