伶盜龍屬
伶盜龍属(屬名:,拉丁文中意为「敏捷的盗贼」)又称盜伶龍屬、快盜龍属,或在中文圈被錯誤翻譯为“迅猛龍”,是蜥臀目兽脚亚目馳龍科恐龍的一屬,大约生活于7,500萬至7,100萬年前的白堊紀中晚期[1],並且伶盜龍和現代鳥類、猛禽之類的關係非常接近。伶盗龍的模式种为蒙古伶盗龍(V. mongoliensis),化石最常发现於蒙古國及中國的內蒙古附近的地区;第二個種為奧氏伶盜龍(V. osmolskae),是在2008年被命名,化石是一個發現於中國內蒙古的頭骨。過去曾經有其他的種,但現多已不被承認。
| 伶盜龍属 化石时期:上白堊紀 75–71 Ma | |
|---|---|
 | |
| 蒙古伶盜龍的骨架模型,位於比利時自然科學博物館 | |
 | |
| 根據已知材料繪製的蒙古伶盜龍重建圖,翅膀重建自化石上的羽莖瘤 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龙总目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 兽脚亚目 Theropoda |
| 科: | †驰龙科 Dromaeosauridae |
| 演化支: | †真驰龙类 Eudromaeosauria |
| 亚科: | †伶盜龍亚科 Velociraptorinae |
| 属: | †伶盜龍属 Velociraptor Osborn, 1924 |
| 种 | |
| |
伶盜龍的體型接近火雞的大小,小於其他的大型馳龍科恐龍(例如恐爪龍),但牠們之間仍有許多相同的生理特徵。伶盜龍是種二足、肉食性的有羽毛恐龍,具有長而堅挺的尾巴,尾巴和手臂上有著如同現代鳥類一樣的羽毛;伶盜龍最明顯的特點是在雙腳的第二腳趾具有大型的鐮刀狀趾爪,這些趾爪被認為用來扎入、刺穿或者撕裂牠們的獵物。伶盜龍與其他馳龍科的差別在於長而低矮的頭顱骨,以及朝上微翹的口鼻部。對於古生物學家而言,伶盜龍是種具有重要研究價值的恐龍,目前已發現超過10個伶盜龍的化石,是馳龍科中數量最多的。而其中一個著名的標本,則保存了與原角龍纏鬥中的伶盜龍。
伶盜龍在大眾流行文化中被錯誤称為“迅猛龍”,是最廣受一般人們熟悉的恐龍之一,這是由於在麥克·克萊頓的小說以及知名電影《侏羅紀公園》(Jurassic Park)中的搶眼表現,並且在續作中《侏羅紀世界》更是對其如人類一般的高智慧有著重描寫,因此有人甚至把《侏羅紀公園》中的“迅猛龍”一詞來代指原本的伶盜龍;但實際上小說與電影版本對於伶盜龍的描述有誤,现实中的伶盜龍只有像貓或者雞一樣大,侏羅紀系列電影中的迅猛龍卻和人類一樣高、有三米長、可輕易捕殺牛或者帶著槍支的武裝人類,與其說是伶盜龍,不如說是“以阿基里斯龍為原型的戰鬥力加強版”可能還更為合適。
簡介

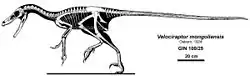
伶盜龍是一种中型驰龍类,成年個體身長估計约2.07公尺,臀部高约0.5公尺,体重推测约80公斤[2]。与其他驰龍类相比,伶盜龍具有相當长的头顱骨,長達25公分;口鼻部向上翘起,使得上側有凹面,下側有凸面。牠們的嘴部有26到28颗牙齒,牙齒间隔寬,牙齒後側有明顯鋸齒邊緣,這特征證明牠們可能是活跃的捕食動物,可以捕食行动迅速的猎物[3][4]。牠們的大脑较大,脑重/体重比在恐龍中相當大,显示牠們是一种非常聪明的恐龍。
类似其他驰龍类,伶盗龍具有大型手部,在結構與靈活性上類似現代鳥類的翅膀骨頭。手部有三根锋利且大幅弯曲的指爪,第二指爪是當中最長的一根,而第一根指爪是最短的。伶盜龍的腕部骨頭結構可以作出往內轉、以及向内抓握的動作,而非向下抓握,非常灵巧[5]。

如同其他的獸腳類恐龍,伶盜龍的第一根腳趾是小型的上爪。但與其它以三根腳趾行走的獸腳類恐龍相比,伶盗龍只依靠后肢的第三、四趾行走,驰龍类都是以這種方式行走。伶盗龍的第二腳趾可以向上、向後收起离开地面,上有大型、鐮刀狀的趾爪,這是牠們著名的重要原因,也是驰龍科與伤齿龍科的典型特徵。這個大型趾爪的外緣長度可达6.5公分,是可怕的攻击武器,可能用來撕開獵物[6][7]。
伶盜龍尾椎上側的前關節突,以及骨化的肌腱,使牠們的尾巴堅挺。前關節突開始於第10節尾椎,往前突出,支撐前面4到10根其他的脊椎,數量依所在位置而定。這些結構使得整個尾巴在垂直方向几乎不能弯曲,但一個伶盜龍標本保存了完整的尾巴骨頭,這些骨頭以S狀水平彎曲,顯示尾巴在水平方向有良好的运动灵活性。这样的尾巴可以帮助伶盗龍在高速奔跑时保持平衡和灵活转向,也说明了伶盗龍是出色的奔跑者[6][7]。
在2007年,古生物學家愛倫·特納(Alan Turner)、彼得·馬克維奇(Peter Makovicky)、馬克·諾瑞爾(Mark Norell)以及他們的同僚宣稱在一個來自於蒙古的伶盜龍化石的前臂發現了羽莖瘤(Quill knobs),確定伶盜龍具有羽毛[8]。


发现歷史
在1922年,美國自然史博物馆的一支探险队在蒙古的戈壁沙漠中发现了第一个伶盗龍的化石标本;該標本(編號AMNH 6515)包含一個遭到壓碎,但是完整的頭顱骨,以及第二趾爪[3]。两年后,该馆的科学家亨利·費爾德·奧斯本(Henry Fairfield Osborn)在确定该标本屬於一種肉食性恐龍後,將牠們命名为蒙古伶盗龍(V. mongoliensis);但奧斯本將該第二趾爪誤認為來自於手部。伶盜龍的屬名衍化自拉丁語,velox意為「快捷的」,raptor意為「強盜」或「盜賊」,意指牠們善於奔跑的身體結構,以及肉食的食性;種名則是發現地蒙古為名[3]。同一年稍早,奧斯本則在大眾媒體上發表了一篇相關文章,並將牠們取名為Ovoraptor djadochtari[9]。但因為Ovoraptor並未在科學文獻與相關的正式文件中被提到,因此該名稱的狀態為無資格名稱,而伶盜龍仍保有命名優先權。
第二次世界大战后的冷戰期間,北美洲的挖掘團隊被蒙古人民革命黨執政的蒙古人民共和國所驅離,而苏联和波兰的探险队与蒙古大学合作发现了许多伶盗龍化石标本,其中最著名的是在1971年由波蘭與蒙古團隊所发现的“搏斗中的恐龍”(編號GIN 100/25),该化石保存了了一只伶盗龍和一只原角龍搏斗的场景[6][10][11]。這個標本被蒙古視為國家級的寶藏,但從2000年起,被外借給紐約市美國自然史博物館,以供一個暫時性的展覽[12]。
冷战後期以來,已有數個中国、美国、加拿大、蒙古等国家科学家所参与的科学考察队在中国和蒙古境内发现了多具伶盗龍化石。在1988到1990年間,一個中國與加拿大所組成的挖掘團隊在中國北部發現了伶盜龍的化石[13]。在1990年,一個美國自然史博物館與蒙古國科學院所組成的挖掘團隊抵達戈壁沙漠,發現了數個保存良好的骨骸[7][14]。其中一個缺乏頭部的標本(編號IGM 100/980),被諾瑞爾等人戲稱為「Ichabodcraniosaurus」,以斷頭谷傳說中的角色Ichabod Crane警探為名[15]。諾瑞爾與馬克維奇推論,這個標本可能屬於蒙古伶盜龍,但由於化石不夠完整,所以無法肯定[7]。
在1999年,中國與比利時組成的挖掘團隊發現一個上頜骨與淚骨,屬於伶盜龍,但不同於蒙古伶盜龍。在2008年,Pascal Godefroit等人將其命名為奧氏伶盜龍(V. osmolskae),種名以波蘭古生物學家哈茲卡·奧斯穆斯卡為名[1]。
發現地點

到目前为止,蒙古伶盗龍的所有标本发现于德加多克塔組(Djadochta Formation)地层中,該地層分布於蒙古國的南戈壁省與中國的內蒙古;另外有一些來自於蒙古國的标本,发现于稍年轻的西戈约特组(Barun Goyot Formation),可能屬於蒙古伶盜龍[16],也有可能是近親屬[17]。这两個地层的年代均為上白垩统坎潘阶,約8300萬年前到7000万年前[18][19]。
在德加多克塔組地層中,幾乎每個著名且多產的挖掘地點都發現了蒙古伶盜龍的化石。蒙古伶盜龍的模式標本是在火焰崖的挖掘地點所發現(該地也名為Bayn Dzak與Shabarakh Usu)[3],而「搏鬥中的恐龍」化石則是在圖格里克(Tugrig)挖掘地點所出土(又名Tugrugeen Shireh)[11]。巴魯恩戈約特組的Khulsan與Khermeen Tsav也是著名的挖掘地點,出土了大量的相關化石,可能屬於伶盜龍[20]。
近年來,在中國內蒙古的巴音满都呼組(Bayan Mandahu Formation)出土了許多幼年的蒙古伶盜龍化石,包含部分骨骼與牙齒。巴音满都呼組的年代與德加多克塔組相同,是一個產量豐富的挖掘地點之一[13]。但是到2008年為止,這些化石還沒有經過處理、清理程序[1]。同樣在2008年,巴音滿都呼組的一個成年個體的部分顱骨被建立為新種,奧氏伶盜龍[1]。
古生態學

這些挖掘地點都處於乾旱的環境中,佈滿沙丘,偶有間歇性的溪流,而巴魯恩戈約特組的環境比年代較古老的德加多克塔組較為濕潤[19]。根據某些標本的死亡姿勢、普遍良好的保存狀態,以及砂岩環境,顯示這些地層的許多動物是被沙塵暴直接掩埋[1]。
在發現伶盜龍化石的地層組,還發現其他恐龍化石,顯示伶盜龍還與這些恐龍共同生存於相同環境。德加多克塔組已發現蒙古伶盜龍、安氏原角龍、谷氏繪龍等化石;巴音滿都呼組則發現奧氏伶盜龍、巨鼻原角龍、魔頭繪龍等化石。這兩個地層組的地理位置相近,而動物群構成相當類似,主要的差異在於種的層面,顯示兩個地層組有動物相的邊界[1]。但目前無法確定造成動物相邊界的原因,也有可能是地質年代的早晚造成這種差異[17]。
跟蒙古伶盜龍發現於相同地層的恐龍包含:傷齒龍科的蜥鳥盜龍、偷蛋龍科的偷蛋龍、馳龍科的大黑天神龍與惡靈龍、傷齒龍科、阿瓦拉慈龍科恐龍;跟奧氏伶盜龍發現於相同地層的恐龍包含:基礎角龍下目的巨嘴龍、偷蛋龍科的曲劍龍、馳龍科的臨河盜龍、傷齒龍科[17]。
分類爭議

伶盜龍在1924年首次被命名時,是被歸類於恐龍中的斑龍科;這是因為該時期的斑龍科與斑龍屬,被當成「未分類物種集中地」,因此許多肉食性恐龍被歸類於該科中,但彼此卻無接近親緣關係[3]。隨者更多恐龍化石的發現,伶盜龍後來被歸類於馳龍科。
伶盜龍目前被歸類於馳龍科的伶盜龍亞科,是較為衍化的一群馳龍科恐龍。在種系發生學中,伶盜龍亞科通常被定義為:馳龍科中,較接近於伶盜龍,而離馳龍較遠的所有物種。但馳龍科的內部分類是經常更改的。在最初建立的時候,伶盜龍亞科只包含伶盜龍一個屬[6]。後來研究則包含了其他屬,通常為恐爪龍與蜥鳥盜龍[21]。一個最近的親緣分支分類法研究,顯示伶盜龍亞科是個單系群,包含:伶盜龍、恐爪龍、白魔龍、以及蜥鳥盜龍(但分類位置未確定)[22]。
在過去,某些馳龍科的屬有時被歸類於伶盜龍屬中,例如平衡恐爪龍(Deinonychus antirrhopus)、藍斯頓氏蜥鳥盜龍(Saurornitholestes langstoni)。因為伶盜龍較早命名,這些種被歸類於伶盜龍屬時,被重新命名為平衡伶盜龍、藍斯頓氏伶盜龍[2]。但到目前為止,伶盜龍中的已承認種僅有蒙古伶盜龍[4][5][23]、以及奧氏伶盜龍(V. osmolskae)[1]。
在2005年新发表的始祖鸟的标本(瑟马普利斯标本)中,发现了保存很好的第二脚趾,類似馳龍科,这可能说明始祖鸟类与伶盗龍有接近的親緣關係[24]。如果这样的親緣關係被进一步证实,因為始祖鳥較早被命名,包括伶盗龍在内的驰龍科很有可能被改歸類於始祖鳥科,屬於鸟纲[25]。目前至少有一位科學家,將馳龍科歸類於始祖鳥科,如果屬實,這將使得伶盜龍成為一種無法飛行的鳥類[5]。
古生物學

獵食行為
发现于1971年的化石标本“搏斗中的恐龍”,保存了伶盗龍和原角龍搏斗的情形,这提供了伶盗龍是活跃的捕食動物以及其捕食方式的直接证据。當這個標本被發現時,過去一度有假設認為這兩隻恐龍是被淹死的[11]。但因為這個標本是在古代沙丘沉積物所發現的,所以目前的看法是這兩隻動物是被掩埋在沙地中的,原因可能是沙丘倒塌,或者是沙塵暴。從兩隻動物的姿態顯示,掩埋過程應該非常快速。原角龍的前肢與後肢都遺失了,可能是被其他食腐動物吞食了[26]。在2011年,科學家比較恐龍、現代鳥類與爬行動物的鞏膜環大小,提出伶盜龍是夜行性動物,而原角龍可能屬於無定時活躍性的動物,覓食、移動行為跟白天黑夜沒有正相關,只休息短暫時間。著名的原角龍、伶盜龍打鬥化石,可能是發生在夜間或光線昏暗的清晨、黃昏[27]。
驰龍科的后肢第二趾有明顯的鐮刀狀趾爪,传统上认为是用于切开猎物身体與挖去內臟的武器[28]。在“搏斗中的恐龍”标本中,伶盗龍的鐮刀狀趾爪嵌入原角龍的喉嚨中,而原角龍的喙嘴則夾住了伶盜龍的右前肢。這顯示伶盜龍可能是用牠们的鐮刀狀趾爪刺穿獵物喉嚨的重要器官来杀死猎物,例如颈静脉、頸动脉,以及气管,而非割開獵物的腹部。伶盗龍爪的鐮刀狀趾爪內側圓滑,並不銳利,并不适合用于切开、刺穿獵物腹部的坚固皮肤和肌肉。然而,目前只有發現鐮刀狀趾爪的骨質部份,這些趾爪在生前應該覆蓋者角質鞘,所以還是有可能具有銳利的邊緣,但無法長期保存銳利狀態,原因是這些趾爪無法後縮以防止被磨損,也無法像貓一樣磨利趾爪[26]。在2005年,BBC的電視節目《恐龍凶面目》(The Truth About Killer Dinosaurs)測試伶盜龍的趾爪是否適合切開。該電視節目製作了一個伶盜龍後肢模型,並將一塊豬腹肉作為測試用的獵物。雖然鐮刀狀趾爪刺穿了測試用的豬腹肉,但無法劃開牠們,顯示伶盜龍的趾爪無法用來割下獵物的內臟。但由於其他科學家沒有參考或重複這個實驗,所以實驗的結果無法被確定。

恐爪龍是另一种和伶盗龍关系密切的恐龍,恐爪龍的化石通常是成群被发现的。此外,恐爪龍的化石偶爾与腱龍(一種大型草食恐龍)一起被发现,这样就说明恐爪龍很可能是团队合作狩猎的捕食動物[29][30]。目前唯一的馳龍科集體行動證據,是個發現於中國的足跡化石,共有六個大型動物所留下,但仍沒有證據顯示牠們有集體獵食行為[31]。不过,虽然在蒙古發現了许多伶盗龍和其他驰龍科的化石,但是没有發現過成群的化石,所以目前還沒有化石證據可以证明伶盜龍是群體狩猎的[23]。伶盗龍也通常被认为是成群狩猎的捕食動物,如同電影《侏羅紀公園》(Jurassic Park)所呈現的,但沒有足夠證據支持馳龍科與伶盜龍有集體獵食行為。馳龍類的集體獵食理論,是根據數個恐爪龍化石圍繞在一個腱龍化石,目前還沒有其他馳龍類被發現類似的化石證據[32]。
在2011年的一項恐爪龍下目趾爪研究,比對不同恐爪龍類、現代鳥類的腳掌結構、趾爪形狀,根據這些現代鳥類的獵食行為,分析這群恐龍的壓制獵物能力。這項研究認為伶盜龍等馳龍類可能以小型獵物為食。研究發現,許多掠食性現代鳥類也有大型第二趾爪,當牠們獵食小型獵物時,會使用腳掌、趾爪將獵物固定在地面上,用體重施壓於獵物,再用鳥喙吞食這些獵物,尤其是鷹科。這項研究推論馳龍類會以類似的方式捕抓、固定獵物,並開始吞食獵物的身體,直到獵物失血過多、失去生理機能。研究發現馳龍類的後肢、腳掌結構最類似鷹,也有大型第二趾爪、類似的腳掌抓取範圍;馳龍類的跗跖骨短、腳掌力量,則比較類似貓頭鷹。伶盜龍的頜部、前肢結構,也符合這項研究的結論。馳龍科的前肢強壯,但極可能覆蓋者羽毛,牠們吞食獵物時可能會將前肢張開,以伺機保持身體的平衡;尾巴也具有平衡身體的功能。馳龍科的頜部結構虛弱,可能主要用來吞食、撕咬獵物的肉體,而非以快速、強壯力量攻擊獵物。研究人員推測,近鳥類可能共同擁有這些演化特徵,而現代鳥類則繼承這些特徵[33]
食腐行為
在2008年,巴音滿都呼組發現一個帶有齒痕的頜部骨頭,週圍環繞者零散的牙齒。2010年公佈了相關研究,頜部骨頭被認為來自於原角龍,零散牙齒被認為來自於伶盜龍[34]。研究提出這隻伶盜龍是在原角龍被其他掠食動物殺死後,吞食其餘的屍體[34][35]。在2012年,一個伶盜龍標本被發現腹部有個長骨頭,來自於神龍翼龍科,這也支持伶盜龍具有食腐行為的理論[36]。
代謝
伶盜龍可能在某種程度上是溫血動物,因為牠們獵食時必須消耗大量的能量。伶盜龍的身體覆蓋者羽毛,而在現代的動物中,具有羽毛或毛皮的動物通常是溫血動物,牠們身上的羽毛或毛皮可以用來隔離熱量。馳龍科與某些早期鳥類的骨頭生長速率,與現代的哺乳類與鳥類相比,顯示牠們具有較為適中的代謝率。紐西蘭的奇異鳥在生理、羽毛型態、骨頭結構、甚至於狹窄的鼻部結構,相當類似馳龍科;而鼻部結構經常是新陳代謝的關鍵指標。奇異鳥是種高度活躍、無法飛行的鳥類,並具有穩定的體溫以及相當低的代謝率,使奇異鳥成為原始鳥類與馳龍科的代謝參考模型[5]。
羽毛

在馳龍科中,比伶盜龍原始的物種通常身體覆蓋者羽毛,並具有完全發展的有羽毛前肢[37]。而伶盜龍的祖先具有羽毛,可能擁有飛行能力,這使得古生物學家認為伶盜龍也具有羽毛,如同許多現代的無法飛行鳥類,仍保有身體上的羽毛。
過去長期以來,古生物學家認為伶盜龍具有羽毛,但沒有證據可以證明。根據2007年9月份的《科學》雜誌,古生物學家愛倫·特納、彼得·馬克維奇、馬克·諾瑞爾,在一個發現自蒙古的伶盜龍化石(編號IGM 100/981,身長1.5公尺,體重15公斤)的前臂,發現了六個羽莖瘤[8]。鳥類骨頭上的的羽莖瘤可用來固定羽毛,而伶盜龍骨頭上的羽莖瘤則明確顯示牠們也具有羽毛[38]。
根據特納等人的說法,並非所有史前鳥類的化石都發現了羽莖瘤,但沒有發現羽莖瘤,不代表這些史前鳥類缺乏羽毛;紅鸛也沒有羽莖瘤,但有羽毛。羽莖瘤的發現顯示伶盜龍擁有羽毛,而且應該是類似現代鳥類翅膀上的羽毛,包含羽軸與羽支所形成的羽片。這些研究人員並提出,伶盜龍的前臂具有14個次要羽毛,而始祖鳥具有至少12個次要羽毛,小盜龍具有18個,脅空鳥龍則具有10個。他們認為這些羽毛數量的不同,代表者這些動物與現代鳥類的差異程度[8]。
特納等人將伶盜龍的羽毛,視為大型、無法飛行的手盜龍類因為體型的增大,而在演化過程中失去羽毛的證據。特納等人並發現,目前的無法飛行鳥類幾乎沒有羽莖瘤,而伶盜龍的羽莖瘤則証明馳龍科的祖先應該可以飛行,但伶盜龍與其它大型的馳龍科後來卻喪失了飛行能力;然而,馳龍科祖先的羽毛也可能具有其他功能,而非用來飛行。對於無法飛行的伶盜龍,牠們的羽毛可能作為展示物用,或孵蛋時覆蓋牠們的蛋巢,或是在上坡奔跑時增加速度用[8]。
病理
一個蒙古伶盜龍的頭顱骨,被發現有兩排平行的穿孔,跟伶盜龍的牙齒大小、間隔相符合。研究人員推論,這代表這隻伶盜龍與其他伶盜龍打鬥時,頭部被咬傷。此外,由於穿孔並沒有事後癒合的痕跡,這隻伶盜龍可能被咬傷後因此死亡[39]。
明星伶盗龍

伶盗龍出现在各式各样的电影和电视节目,例如《侏羅紀公園》系列電影,以及種種科普片当中。探索频道的纪录片《恐龍星球》(Dinosaur Planet)细致的记述了一只雌性伶盗龍的故事。在BBC的《与恐龍同行》(Walking with Dinosaurs)特别节目《鐮刀龍探秘》(The Giant Claw)中也有伶盗龍出场, 而BBC的《恐龍兇面目》(The Truth About Killer Dinosaurs)中专题介绍了伶盗龍与原角龍的搏斗。
伶盗龍还出现在其他领域如音乐和体育中。例如,NBA球队多伦多猛龍(Toronto Raptors)是以伶盗龍命名,並曾经使用一只伶盗龍作为队徽。在2005年,曾经举行过一次名为“Velociraptour”的演唱会。一个叫Roboraptor的玩具创意也来自伶盗龍。
伶盗龍还成为一些电子游戏的主角,包括《恐龍危机》(Dino Crisis)系列,《恐龍猎人》(Turok)系列和《魔兽争霸》。
參考資料
- (英文)Godefroit, Pascal; Currie, Philip J.; Li Hong; Shang Chang Yong; and Dong Zhi-ming. . Journal of Vertebrate Paleontology. 2008, 28 (2): 432–438.
- (英文)Paul, Gregory S. . New York: Simon and Schuster. 1988: 464pp. ISBN 978-0671619466.
- (英文)Osborn, Henry F. . American Museum Novitates. 1924a, 144: 1–12 [2007-10-08]. (原始内容存档于2011-06-29).
- (英文)Barsbold, Rinchen; Osmólska, Halszka. . Acta Palaeontologica Polonica. 1999, 44 (2): 189–219 [2012-07-03]. (原始内容存档于2009-01-07).
- (英文)Paul, Gregory S. . Baltimore: Johns Hopkins University Press. 2002: 472pp. ISBN 978-0801867637.
- (英文)Barsbold, Rinchen. . Transactions of the Joint Soviet-Mongolian Paleontological Expedition. 1983, 19: 5–119.
- (英文)Norell, Mark A.; & Makovicky, Peter J. . American Museum Novitates. 1999, 3282: 1–45.
- (英文)Turner, A.H., Makovicky, P.J., and Norell, M.A. (2007). "Feather quill knobs in the dinosaur Velociraptor." 页面存档备份,存于 Science, 317(5845): 1721.
- (英文)Osborn, Henry F. . Natural History. 1924b, 24: 133–149.
- (英文)Kielan-Jaworowska, Zofia; & Rinchen Barsbold. . Paleontologica Polonica. 1972, 27: 5–13.
- (英文)Barsbold, Rinchen. . Paleontologica Polonica. 1974, 30: 5–22.
- (英文)American Museum of Natural History. . [2007-06-13]. (原始内容存档于2010-11-23).
- (英文)Jerzykiewicz, Tomasz; Phil Currie; Eberth, David A.; Johnston, P.A.; Koster, E.H.; & Zheng J. . Canadian Journal of Earth Sciences. 1993, 30 (10): 2180–2195 [2012-07-03]. doi:10.1139/e93-190. (原始内容存档于2017-11-11). [printed early 1994]
- (英文)Norell, Mark A.; & Makovicky, Peter J. . American Museum Novitates. 1997, 3215: 1–28 [2007-10-08]. (原始内容存档于2020-02-09).
- (英文)Novacek, M. (1996). Dinosaurs of the Flaming Cliffs, Anchor Books.
- (英文)Weishampel, David B.; Barrett, Paul M., Rodolfo Coria, Le Loueff, Jean, Xu, Xing; Zhao, Xijin; Sahni, Ashok, Gomani, Emily M.P. & Noto, Christopher N. . David B. Weishampel, & Osmólska, Halska (eds.). (编). Second Edition. Berkeley: University of California Press. 2004: Pp. 517–606. ISBN 0520242092.
- (英文)Nicholas R. Longrich, Philip J. Currie, Dong Zhi-Ming. . Palaeontology. 2010, 53 (5): 945–960. doi:10.1111/j.1475-4983.2010.00968.x.
- (英文)Gradstein, Felix M.; Ogg, James G.; & Smith, Alan G. . Cambridge: Cambridge University Press. 2005: 500pp. ISBN 978-0521781428.
- (英文)Jerzykiewicz, Tomasz; & Dale Russell. . Cretaceous Research. 1991, 12 (4): 345–377. doi:10.1016/0195-6671(91)90015-5.
- (英文)Osmólska, Halska. . Phil Currie & Padian, Kevin (eds.). (编). . San Diego: Academic Press. 1997: p.41. ISBN 0-12-226810-5.
- (英文)Currie, Philip J. . Journal of Vertebrate Paleontology. 1995, 15 (3): 576–591 [2007-10-29]. (原始内容存档于2007-11-17).
- (英文)Norell, Mark A.; Clark, James M.; Turner, Alan H.; Makovicky, Peter J., Rinchen Barsbold; & Rowe, Timothy. . American Museum Novitates. 2006, 3545: 1–51 [2007-10-29]. (原始内容存档于2020-03-12).
- (英文)Norell, Mark A.; & Makovicky, Peter J. . David B. Weishampel, Peter Dodson & Osmólska, Halska (eds.). (编). Second Edition. Berkeley: University of California Press. 2004: Pp. 196–209. ISBN 0520242092.
- (英文)Mayr, G., Pohl, B., & Peters, D.S. 2005. A well-preserved Archaeopteryx specimen with theropod features. Science, 310: 1483-1486. doi:10.1126/science.1120331.
- (英文)Sereno, P.C., McAllister, S., & Brusatte, S.L. 2005. TaxonSearch 页面存档备份,存于: a relational database for suprageneric taxa and phylogenetic definitions. PhyloInformatics 8: 1-21.
- (英文)Carpenter, Kenneth. (PDF). Gaia. 1998, 15: 135–144 [2012-07-03]. (原始内容 (PDF)存档于2006-01-11).
- (英文)Schmitz, L.; Motani, R. . Science. 2011, 332 (6030): 705–8. PMID 21493820. doi:10.1126/science.1200043.
- (英文)Ostrom, John H. . Bulletin of the Peabody Museum of Natural History. 1969, 30: 1–165.
- (英文)Maxwell, W. Desmond; & John Ostrom. . Journal of Vertebrate Paleontology. 1995, 15 (4): 707–712 [2007-10-29]. (原始内容存档于2007-09-27).
- (英文)Brinkman, Donald L.; Cifelli, Richard L.; & Czaplewski, N.J. . Oklahoma Geological Survey Bulletin. 1998, 146: 1–27.
- (英文)Li, Rihui; Lockley, M.G., Makovicky, P.J., Matsukawa, M., Norell, M.A., Harris, J.D. and Liu, M. . Naturwissenschaften. 2007. (原始内容存档于2020-02-09).
- (英文)Long, John (2008). Feathered Dinosaurs: The Origin of Birds. Oxford University Press. ISBN 978-0-19-537266-3, p. 21.
- (英文)Fowler, D.W.; Freedman, E.A.; Scannella, J.B.; Kambic, R.E. . PLoS ONE. 2011, 6 (12): e28964 [2012-07-03]. PMID 22194962. doi:10.1371/journal.pone.0028964. (原始内容存档于2015-01-02).
- (英文)Hone, David; Choiniere, Jonah; Sullivan, Corwin; Xu, Xing; Pittman, Michael; Tan, Qingwei. . Palaeogeography, Palaeoclimatology, Palaeoecology. 2010, 291 (3–4): 488−492. doi:10.1016/j.palaeo.2010.03.028.
- (英文)Walker, Matt. . BBC Earth News. 2010-04-06 [2010-08-20]. (原始内容存档于2018-06-12).
- (英文)Hone, D.; Tsuihiji, T.; Watabe, M.; Tsogtbaatr, K. (2012). "Pterosaurs as a food source for small dromaeosaurs". Palaeogeography, Palaeoclimatology, Palaeoecology 331-332: 27. doi:10.1016/j.palaeo.2012.02.021
- (英文)Xu, Xing; Zhou, Zhonghe; Wang, Xiaolin; Kuang, Xuewen; Zhang, Fucheng; Du, Xiangke. . Nature. 2003, 421 (6921): 335–340. PMID 12540892. doi:10.1038/nature01342.
- (英文)Turner, Alan. . Nature. 2007 [2007-10-29]. (原始内容存档于2019-02-22).
- (英文)Molnar, R. E., 2001, Theropod paleopathology: a literature survey: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, p. 337-363.
外部链接
| 维基共享资源中相关的多媒体资源:伶盜龍 |
| 维基语录上的相关摘錄: Velociraptor |
| 維基教科書中的相關電子:Wikijunior Dinosaurs/Velociraptor |
- American Museum of Natural History. "Fighting Dinosaurs: New Discoveries from Mongolia: Videos." c. 2000. Three videos related to a fight between Protoceratops and Velociraptor.
- Hartman, Scott. "Velociraptor." SkeletalDrawing.com. Several artistic renditions of Velociraptor.
- Western Australian Museum – Dinosaur Discovery – Fight to the death 页面存档备份,存于