真核起始因子2
eIF2(eukaryotic Initiation Factor 2,真核起始因子2)是一个重要的真核起始因子,它的作用是在真核翻译起始过程中介导起始tRNA(Met-tRNAi)与核糖体结合。它是一个异源三聚体,由α、β、γ三个亚基组成。eIF2也是第一个被发现的其蛋白磷酸化作用能够调控真核翻译的起始因子。
功能
eIF2在真核翻译起始过程中扮演着关键角色。它与GTP以及起始tRNA形成三聚体复合物(TC),在eIF3和eIF5的作用下,TC结合到40S核糖体亚基上共同形成43S前起始复合物。在60S核糖体亚基结合上来之前,eIF5激活了eIF2的GTP酶活性,从而将结合的GTP水解为GDP。eIF2·GDP就从40S核糖体亚基上被释放出来。游离的eIF2·GDP在鸟苷酸交换因子eIF2B的作用下,被重新转换为eIF2·GTP从而可以再次参与起始过程。因此,eIF2的活性在细胞内可以被eIF2B调节,eIF2B的作用是使足量的eIF2·GTP存在以参与起始过程。[1][2][3][4]
亚基组成和结构
eIF2是一个异源三聚体,由α、β、γ三个亚基组成。三个亚基的氨基酸序列高度保守(人与酵母对应亚基之间的序列等同性高达47 – 72 %)。
| 亚基 | α | β | γ |
|---|---|---|---|
| 分子量/kDa | 36 | 38 | 52 |
| 相似性 | eIF2α家族 | eIF2β/eIF5家族 | GTP结合的延伸因子家族 |
| 相互作用 | 结合eIF5、eIF2B和RNA | 结合GTP和RNA |
α亚基包含两个球形结构域,N端结构域和C端结构域,这两个结构域之间几乎没有相互作用,其各自都可以独立正确折叠。[6]α亚基被认为是eIF2中的调节亚基。它是磷酸化调控的主要对象,即对位于51位的丝氨酸(Ser51)进行磷酸化,而α亚基的磷酸化形式能够调节eIF2B的活性。N端结构域上还含有一个S1亚结构域,可能为RNA结合位点。
β亚基也含有多个磷酸化位点(2, 13, 67, 218位上的氨基酸残基)。它的N端结构域还含有3个赖氨酸聚集的区域,是与eIF5结合的重要位点。C端结构域含有eIF2B的结合区和一个预测的锌指花样区,其中锌指花样参与了TC和43S前起始复合物的形成。它还含有两个保守的鸟苷酸结合区,但不参与eIF2的活性的调控。β亚基还被认为能够结合tRNA和mRNA。
γ亚基含有属于GTP酶,其作用是结合GTP和Met-tRNAi,并在eIF5的激活下,将eIF2结合的GTP水解为GDP。它含有三个保守的鸟苷酸结合区,是GTP/GDP的主要结合位点。它还含有一个tRNA结合凹槽和Zn2+离子结合位点。[3][7][6]
调控
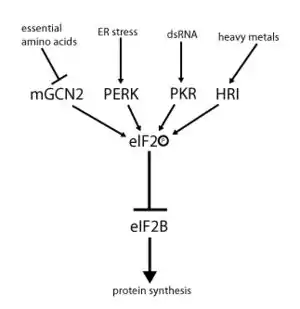
eIF2的活性是通过鸟苷酸交换和磷酸化来共同调控的。磷酸化发生在α亚基中的Ser51,其磷酸化形式表示为eIF2αP,哺乳动物中存在四种不同的激酶来对Ser51进行磷酸化。这些激酶的激活能够应答多种不同的细胞压力和刺激。例如,HRI(heme-regulated inhibitor Kinase)激酶在低血红素、亚砷酸盐中毒或热休克条件下激活,GCN2(generalcontrol non-derepressible-2)激酶在氨基酸缺乏或紫外线损伤下激活,PKR(protein kinase RNA)激酶能被病毒双链RNA激活,PERK(PKR-like endoplasmic reticulum kinase)激酶则能应答内质网压力。一旦被磷酸化,eIF2表现出增强的对eIF2B活性的抑制。由于在细胞中,eIF2的含量要远远高于eIF2B的含量,因此即使是低水平的eIF2α磷酸化也已经可以有效的抑制eIF2B的活性。由于eIF2B是唯一能够对eIF2发挥鸟苷酸交换作用的因子,而eIF2在翻译起始中发挥着关键作用,因此eIF2B活性的降低将会抑制翻译起始。综上所述,通过鸟苷酸交换和磷酸化对eIF2的活性进行调控,也就能够对细胞中整体蛋白翻译水平进行调节。[1][2][3][8]
eIF2的作用机制在真核生物中是相当保守的,它的活性的降低将导致整体蛋白翻译水平的降低,而这种降低可能是细胞对各种应激反应的一种应答方式。此外,eIF2活性的降低将会影响某些转录调节蛋白的翻译,从而从转录水平上影响很多基因的表达。
相关疾病
由于eIF2在翻译起始中发挥着关键作用,因此eIF2上出现缺陷将是致命的。虽然没有发现任何疾病与eIF2上的突变有直接关系,许多疾病却由eIF2活性下调(通过其上游的激酶)所引发。例如,在患有神经退行性疾病(如阿兹海默病、帕金森氏症和亨丁頓舞蹈症)的病人体内发现浓度增加的活化的PKR和被抑制的(即被磷酸化的)eIF2。也有一些疾病是与eIF2的鸟苷酸交换因子eIF2B相关。如白质消失(leukoencephalopathy)[3][9]
参考资料
- (英文)Kimball SR. . Int. J. Biochem. Cell Biol. January 1999, 31 (1): 25–9. PMID 10216940.
- (英文)Hershey JW. . J. Biol. Chem. December 1989, 264 (35): 20823–6. PMID 2687263.
- (英文)Hinnebusch AG. . Annu. Rev. Microbiol. 2005, 59: 407–50. PMID 16153175. doi:10.1146/annurev.micro.59.031805.133833.
- (英文)Proud CG. . Semin Cell Dev Biol. 2005, 16: 3–12.
- (英文)Kimball SR, Jefferson LS. . Nutr Metab (Lond). 2004, 1 (1): 3. PMC 524028. PMID 15507151. doi:10.1186/1743-7075-1-3.
- (英文)Ito T, Marintchev A, Wagner G. . Structure. September 2004, 12 (9): 1693–704. PMID 15341733. doi:10.1016/j.str.2004.07.010.
- (英文)Roll-Mecak A, Alone P, Cao C, Dever TE, Burley SK. . J. Biol. Chem. March 2004, 279 (11): 10634–42. PMID 14688270. doi:10.1074/jbc.M310418200.
- (英文)Nika J, Rippel S, Hannig EM. . J. Biol. Chem. January 2001, 276 (2): 1051–6. PMID 11042214. doi:10.1074/jbc.M007398200.
- (英文)Chang RC, Yu MS, Lai CS. . Neurosignals. 2006, 15 (5): 249–58. PMID 17496426. doi:10.1159/000102599.
外部链接
- MeSH(醫學主題詞)上面的EIF-2(美式英语)