酵母
酵母(拼音:中國大陆:jiàomǔ、台灣:xiàomǔ;注音:中國大陆:ㄐㄧㄠˋ ㄇㄨˇ、台灣:ㄒㄧㄠˋ ㄇㄨˇ;英文:Yeast)是真核生物域真菌界之下的一個非分类学术语,泛指能发酵糖類的各种单细胞微生物。不同的酵母菌在进化和分类地位上有异源性;而酵母菌种类很多,已知的约有56属500多种[1]。一些酵母菌能夠通過出芽的方式進行無性生殖,也可以通過形成孢子的形式進行有性生殖。酵母經常被用於酒精釀造或者麵包烘培行業。目前已知有1500多種酵母,大部分被分類到子囊菌門。[2]酵母菌屬兼性厭氧菌。[3]
| 酵母菌 | |
|---|---|
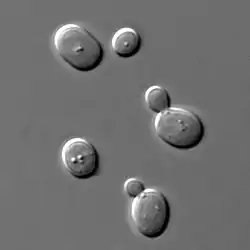 | |
| 酿酒酵母 | |
.png.webp) | |
| 典型酵母细胞的横截面示意图 | |
| 科学分类 | |
| 总域: | 新壁總域 Neomura |
| 域: | 真核域 Eukaryota |
| 演化支: | 單鞭毛生物 Unikonta |
| 演化支: | 後鞭毛生物 Opisthokonta |
| 总界: | 菌物总界 Holomycota |
| 界: | 真菌界 Fungi |
| 門與亞門 | |
担子菌门 Basidiomycota p. p. | |
自從第一種酵母於數億年前出現,現時已辨識出1,500個酵母的物种[4][5][6],佔所有已描述的真菌物種的1%[7]。酵母是從多細胞生物演化成的单细胞生物[8],有些物種可以長成多細胞的假菌絲[9]。不同酵母在不同環境中的體積差異很大,通常直徑在3~4微米,但有些可以長到40微米[10]。大部份的酵母透過有絲分裂行無性生殖,有些在分裂時兩個後代的體積不對稱,稱為出芽生殖。
歷史
4000年前,古埃及人已經開始利用酵母釀酒與製作麵包了[11] ;中國的殷商時期(約3500年前),古人利用酵母釀造米酒,而酵母饅頭、餅等開始於漢朝時期。[5]
1680年,荷蘭科學家安東尼·范·列文虎克首次利用顯微鏡觀察到酵母,但當時並沒有將其當作一個生物體看待。[12]
1857年,法國科學家路易·巴斯德首次發現釀造酒精來自酵母體的發酵作用,而並非簡單的化學催化。[13][14]巴斯德曾經將空氣通進釀酒液中,發現酵母的細胞量增加了,但是酒精的生成量減少,後來人們將此現象稱為“巴斯德效應”。[15]
酵母的工業化生產與商業化依賴於乾燥與壓濾技術的發展。1846年,歐洲實現酵母的工業化生產。[5]美國酵母的工業與商業化是隨著1876年費城百年博覽會的舉辦展開的。中國酵母的現代化生產開始於20世纪80年代中期。[5]
細胞形態與結構
酵母細胞明顯比大多數細菌大,細胞大小約為 2~5 × 5~30μm(短軸×長軸)。[16]酵母多數為單細胞生物,常呈卵圓形或者圓柱形。[17]實際上,每種酵母確實具有自己特有的形態模式,但會隨著菌齡與環境不斷變化。[18]一般平板培養基上的酵母菌落呈白色凸起粒狀,常帶有酒香味。
酵母属于真核微生物,除没有鞭毛外[19],一般都具有细胞壁、细胞膜、线粒体、核糖体、液泡等细胞器。
- 細胞壁:厚度為0.1~0.3μm,不如細菌的堅韌;主要成分為葡聚醣、甘露聚醣等。[16]酵母細胞壁呈“三明治”形:內層葡聚醣、外層甘露聚醣以及中間蛋白層。[20]有研究表明,葡聚醣是維持細胞壁內壁強度最主要的物質。[21]
- 細胞膜:細胞膜為磷脂雙分子層,與其他生物一樣都是雙膜中間鑲嵌著蛋白質。此外,酵母細胞膜中還含有甾醇,其中以麥角甾醇最為常見。[16]

营养与生长
酵母菌廣泛生活於潮濕且富含糖分的物體表層,例如果皮表層、土壤、植物表面、植物分泌物(如仙人掌的汁),甚至空氣中也有分佈。此外,有研究發現酵母還能寄生於人類身上與一些昆蟲腸道內。[23]
酵母菌屬於化能異養、兼性厭氧型微生物,能夠直接吸收利用多種單醣分子,比如葡萄糖、果糖等。一些酵母菌還能代謝利用五碳糖[24]、乙醇或者有机酸。一部分双糖,例如蔗糖,能在胞外酶作用下水解为单糖被吸收利用。[25]酵母菌不能直接利用淀粉等多糖类物质。因此,在啤酒酿制过程中,原料麦必须经过糖化才能被酿酒酵母进一步发酵利用。[25]
許多酵母營專性或兼性好氧的生活方式,目前尚未發現專性厭氧的酵母。在缺乏氧氣時,發酵型的酵母會進行缺氧呼吸作用,當中通過糖酵解作用將葡萄糖轉化成丙酮酸,其後丙酮酸經脫碳作用脫去碳原子,形成乙醛,同時釋出CO2,乙醛再被於糖酵解作用產生的NADH2還原成乙醇並產生能量(ATP)。
- C6H12O6 →2C2H5OH + 2CO2 + 2ATP
在釀酒過程中,乙醇被保留下來;在烤麵包或蒸饅頭的過程中,CO2將麵團發起,而酒精則揮發。在有氧條件下,酵母將葡萄糖經有氧呼吸(糖酵解→三羧酸循環)代謝生成CO2和H2O。
- C6H12O6 + 6O2→6CO2 + 6H2O + 30(32)ATP
有氧條件下,酵母菌往往能夠迅速出芽繁殖。
酵母菌的最適生長溫度各異,在自然pH或弱酸環境中生長生活力最高。畢竟酵母能在低pH(pH≈3)條件下生長。[26]
繁殖
酵母具有无性繁殖和有性繁殖两种方式。
无性繁殖
- 出芽生殖(budding) 在生长环境良好时,酵母菌迅速生长,几乎每个细胞外面都会产生芽体,而且芽体上会产生新的芽体。芽体逐渐长大成熟后与母体分离。
- 分裂生殖(fission) 少数酵母菌如裂殖酵母属(Schizosaccharomyces)具有与细菌一样的二分裂繁殖方式。
- 孢子生殖 部分少数酵母菌如掷孢酵母属(Sporobolomyces)能在其营养细胞上长出小梗,小梗上产生掷孢子。孢子成熟后,通过一种特有的喷射机制将孢子喷出。[27]
用途
酵母具备许多诱人的特征,广泛应用于工业、商品生产、环保以及科学研究领域。酿造酒精与面包烘培是酵母菌最常见、最古老的利用方式。此外, 许多酵母还能用于生产各类饲料以及工业营养物,比如SCP(Single Cell Protein)、酵母提取物等。某些酵母耐酸、耐高渗透、分解吸收有毒物质,同时被广泛应用于污水处理领域。[28]在科学研究上,酿酒酵母(Saccharomyces cerevisiae)作为模式生物被使用[29];另外一些酵母已经被开发为异源蛋白表达系统使用,利用基因技术在酵母细胞内表达外源蛋白质。
酒精饮料
酵母菌被广泛应用于酒精饮料,例如啤酒、果酒、蒸馏酒的生产中,酵母菌在无氧条件或低氧浓度条件下,消耗谷物、水果等碳水化合物原料,为自身提供能量并产生酒精与二氧化碳。最常见的用于啤酒与果酒酿造的菌种为酿酒酵母(Saccharomyces cerevisiae)。[30]
啤酒

用于酿造啤酒的酵母菌,根据发酵类型的不同,主要分为两大类:爱尔酵母(ale yeast)与拉格酵母(窖藏酵母)(lager yeast)。[30]爱尔酵母发酵期间会慢慢上升至啤酒表层,因此又称顶層发酵酵母(top fermenting yeast)。最常用的爱尔酵母为啤酒酵母(Saccharomyces cerevisiae)。[31] 由爱尔酵母发酵的啤酒有:爱尔啤酒、麦啤、司陶特(stouts)等。
拉格酵母(窖藏酵母)用于底层发酵(bottom fermentation)。与顶層发酵方法相比,底层发酵往往采用较低的发酵温度,发酵时间较长。到发酵末期,酵母菌下沉于酒桶底部,由此啤酒酒色也较为透明。[30]卡尔斯博酵母(Saccharomyces pastorianus, 舊稱Saccharomyces carlsbergensis)是一种典型与比较常用的拉格酵母(窖藏酵母)之一。现在,爱尔酵母与拉格酵母(窖藏酵母)已被重新归类于S. cerevisae菌属。[31]
此外,还有许多种类的酵母菌应用在酒精酿制中,以适应不同工艺与口感风味上的需要。目前。各种各样的育种技术被引进到优良菌种的选育中;基因工程菌技术的加入,赋予了酵母菌自然菌种所不具备的新特性。[32]有研究称,转入黑曲霉菌葡萄糖淀粉酶基因的酵母工程菌,能够更高效的分解利用原来中的淀粉。[33]
葡萄酒

传统葡萄酒的酿造,便是利用粘附于果皮上的天然酵母菌来酿制,此方法亦成为自然发酵法。这些果皮上的菌种,其实是许多微生物的“混合体”,某种程度上可认为其增加了酿酒过程及产品质量的许多不确定因素。因此,现在越来越多的酿酒师和酿酒厂会选择经分离纯化后的纯菌种进行发酵。[34]
原因是當某些特定種類的細菌(如葡萄球菌 Staphyloccus gallinarium)也存在於培養液中的時候,這些細菌會透過跟酵母菌的溝通與互動,使得酵母菌的[GAR+]普利昂蛋白被引發出來;導致酵母菌就不再專心利用葡萄糖,轉而利用其他碳水化合物。而當酵母菌不再專心利用葡萄糖時,發酵作用就不會大量發生,導致培養液內的酒精濃度也不會像正常發酵進行時的快速上昇,也不會提高到細菌無法忍受的濃度。這麼一來,只是讓細菌得到一個較好的生長環境,而酵母菌還是可以生長,,而且[GAR+]的酵母菌,比原來的酵母菌更能夠耐酒精,也更能夠在養分不足(低氨基酸)的環境下生存,但這不會是釀酒廠所樂見的事。除了將酵母菌純化之外,也可以在壓碎葡萄時就加入二氧化硫(SO2,葡萄酒釀造中最常使用的殺菌藥劑,通常在發酵前以及發酵結束後加入)來殺菌,降低其他的細菌與酵母菌「聊天」的機會,便可減少發酵停滯現象的發生[35]。
葡萄酒風味的多元也跟酵母菌的多元有關,不同的酵母菌,可形成不同风味的葡萄酒。
致病性
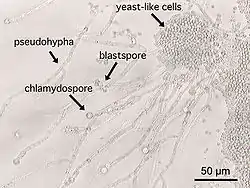
一般酵母菌被指认为是一种条件致病菌,特别容易对免疫力低下的病人造成感染。酵母菌感染属于真菌感染中的一种形式。
白色念珠菌(Candida albicans)能够引起鹅口疮以及尿道炎等感染疾病。白色念珠菌在人类身上主要出现在口腔、肠道、尿道等部位的粘膜上,小部分生活在皮肤表面。正常情况下,念珠菌以酵母细胞型存在,没有致病性;在一些因素的诱导下,比如免疫力缺陷、过量使用抗生素等,白色念珠菌大量转化为菌丝生长型,并大量繁殖,入侵患者粘膜系统,引起炎症而发病。在怀孕晚期服用避孕药的妇女中,极易感染尿道炎,其中一个可能的诱因便是身体上的激素出现了失衡。[37]
白色隐球菌(Cryptococcus albidus)是一种一般对人类无害的出芽型酵母菌。但在免疫系统缺陷者身上,可能感染病人引起一种名为隐球菌病(cryptococcosis)的疾病。[38] 另外,有案例显示,一位进行免疫抑制治疗的病人肺部受到白色隐球菌的感染后,导致出现急性呼吸窘迫综合症(ARDS)的病症。[39]
酿酒酵母(Saccharomyces sereviciae)一般不被认为是条件性致病菌,但是也有少量的报告显示出酿酒酵母具有致病的能力。[40][41]
参考文献
- 车振明. [ ] 请检查
|url=值 (帮助). : 科学出版社. 2011年6月: 55頁 [ ]. ISBN 978-7-03-031216-7 (中文(简体)). - . ARS. [2014-09-01]. (原始内容存档于2008-02-22) (英语).
- Müller, Gunther. . Magdeburg, Deutschland: VerlagsKG Wolf. 2011: 3-6. ISBN 978-3-89432-707-1 (德语).
- (英语).:The second completely sequenced yeast genome came 6 years later from the fission yeast Schizosaccharomyces pombe, which diverged from S. cerevisiae probably more than 300 million years ago.
- E5%8E%86%E5%8F%B2-history-of-yeasts/ 请检查
|url=值 (帮助). [2011-08-17]. (原始内容存档于2019-06-14). - Hoffman CS, Wood V, Fantes PA. . Genetics. 2015-10, 201 (2): 403–23. PMC 4596657. PMID 26447128. doi:10.1534/genetics.115.181503 (英语).
- (德语).
- Huxley A. . Collected Essays. 1871 [2009-11-28]. (原始内容存档于2011-09-07).
- Phillips T. . Science@NASA. [2009-11-28]. (原始内容存档于2009-11-04).
- Barnett JA. . Microbiology (Reading, Engl.). 2003, 149 (Pt 3): 557–67 [2009-11-28]. PMID 12634325.
- .
- 岑沛霖, 蔡謹. . 北京, 中國: 化學工業出版社. 2000年6月: 72. ISBN 7-5025-2643-9.
- 杨非,曹萌,金怡,杨秀山,田沈. . 生物工程学报. 2012, 28 (8): 901–911.
- Tintelnot K. . Der Pathologe. 2013, 34 (6): 519–527. doi:10.1007/s00292-013-1828-4.
- Prescott, Harley, Klein. . Hammond, IN, U.S.A.: McGraw-Hill Higher Education. 2002-10-25: 554. ISBN 0-07-282905-2.
- 周德慶. . 北京.中國: 高等教育出版社. 2002年5月: 47-50. ISBN 978-7-04-011116-3.
- . [2014-08-31]. (原始内容存档于2014-09-03) (中文(简体)).
位于内层的葡聚糖是维持细胞壁强度的主要物质。
- . (原始内容存档于2015-02-05) (中文(简体)).
- Suh SO, McH. JV., Pollock DD., Blackwell M. . Mycol Res. 2005, 109 (Pt 3): 261–265 [2011-10-19]. PMID 2943959.
- Barnett JA. . Journal of General Microbiology. 1975, 90 (1): 1–12. PMID 1176959.
- . Howstuffworks. [2011-10-19].
Other yeast enzymes can make simple sugars out of disaccharides (double sugars), which are found in certain organisms.
- . Invitrogen. [2011-10-19]. (原始内容存档于2019-06-14).
- 周德青. 第二版. 北京: 高等教育出版社. 2005年5月: p57–62. ISBN 7040111160.
- 曹文平, 武晓刚, 郭., etc. . 中国生物工程杂志. 2007, 27 (11): 99–104 [2011-08-25].
- . Lallemand. 2011-04-27 [2011-08-25]. (原始内容存档于2011-09-26).
- . University of Hawai`i at Manoa. 2011-3 [2011-08-27]. (原始内容存档于2009-11-30) (英语).
This, along with modifications in the brewing process have led to different types of beers. Those most often seen in North America include: ...
- . BeerAdvocate. [2011-08-27] (英语).
- . 杯莫停_beimoting.com. [2011-08-27]. (原始内容存档于2013-07-29) (中文).
- 程少菊, 张添元, 罗进贤 等. . 中山大学学报《自然科学版》. 2007, * (3) [2011-08-27].
将酿酒酵母的rDNA片段,黑曲霉葡萄糖淀粉酶基因表达盒及G418抗性基因表达盒重组进经过改造的质粒pSP72,构建酿酒酵母整合型质粒YIp4RGAn及YIp19RGAn,转化酿酒酵母实验室菌株GRF18、生产菌株JL108、SD和JM,获得能高效表达葡萄糖淀粉酶和分解淀粉的酿酒酵母基因工菌.
- 王江松 许引虎 安琪酵母股份有限公司. . FAM365.com. 2009-08-04 [2011-08-27] (中文).
- {Daniel F. Jarosz, Jessica C.S. Brown, Gordon A. Walker, Manoshi S. Datta, W. Lloyd Ung, Alex K. Lancaster, Assaf Rotem, Amelia Chang, Gregory A. Newby, David A. Weitz, Linda F. Bissonemail, Susan Lindquist. 2014. Cross-Kingdom Chemical Communication Drives a Heritable, Mutually Beneficial Prion-Based Transformation of Metabolism. Cell. 158(5):1083–1093.}
- 福王寺明. . 台北,台灣: 台灣東販. 2009年1月: 5-20. ISBN 978-9-86-176844-1 (中文(繁體)).
- Jim Deacon, Institute of Cell and Molecular Biology, The University of Edinburgh. . [2011-09-23]. (原始内容存档于2006-09-25) (英语).
- Jim Deacon, Institute of Cell and Molecular Biology, The University of Edinburgh. . [2011-09-23]. (原始内容存档于2006-09-25) (英语).
C. neoformans, a significant pathogen of immunocompromised people, causing the disease termed cryptococcosis.
- Burnik C, Altintas ND, Ozkaya G, Serter T, Selçuk ZT, Firat P, Arikan S, Cuenca-Estrella M, Topeli A. . Medical mycology. 2007, 45 (5): 469–73. PMID 17654275.
- J K Byron, K V Clemons, J H McCusker, R W Davis, and D A Stevens. . Infection and Immunity. 1995, 63 (2): 478–85. PMC 173020. PMID 7822013.
- . TSCA Biotechnology. 1997-2 [2011-09-23] (英语).
- . Alimentary Pharmacology & Therapeutics. 2007-09-15 [2014-06-30] (英语).
