致病真菌
致病真菌(英語:Pathogenic Fungi)是引起人類或其他生物產生真菌病的真菌。 已知大約300種真菌對人類具有致病性[1]。雖然真菌是真核生物,但是許多致病真菌也是微生物[2]。對人類致病的真菌的研究稱為醫用真菌學(medical mycology),對植物致病的真菌和其他生物的研究被稱為植物病理學。
假絲酵母屬
假絲酵母屬的菌種是重要的人體病源,已確知會造成免疫缺陷的宿主(如接受器官移植者、AIDS患者、癌症患者)的機會性感染。感染難以治療且可能非常嚴重:30~40%的系統性感染會造成死亡。白色念珠菌和其他數種在醫學上重要的假絲酵母屬菌種的基因體已完成定序,使假絲酵母屬的比較和功能性基因體分析的研究變得可行。這些研究助於發展敏感性診斷策略和新的抗真菌療法。[3]
麴菌屬

形成氣熔膠的麴菌屬孢子幾乎到處都找得到,所以人們經常持續地暴露於這些孢子。此種暴露是正常人體狀態的一部分且一般不會損害健康。然而,麴菌可經由三種主要方式致病:產生真菌毒素、引起過敏反應與局部或系統性感染。後兩種致病方式主要受宿主免疫狀態影響,由宿主主動免疫反應對抗真菌孢子或菌絲所致。相較之下,若免疫系統已經瓦解、抵抗力微弱或缺乏,可能感染嚴重的麴菌症[4]。
最常見致病菌種是煙麴黴菌和黃麴黴菌。黃麴黴菌產生的黃麴毒素是毒物也是致癌物,可能污染食物如堅果。煙麴黴菌和棒麴黴菌可造成過敏疾病。某些麴菌屬菌種可造成穀類作物疾病,尤其是玉蜀黍,並合成黃麴毒素等黴菌毒素。麴菌症是麴菌屬引起的一組疾病。症狀有發燒、咳嗽、胸痛、呼吸急促。通常只有免疫系統衰弱或其他肺部疾病的患者才容易感染。[2][4]
煙曲霉(Aspergillus fumigatus)的孢子在大氣中普遍存在。煙曲霉是一種機會性致病菌。它可能在免疫功能低下的個體中引起潛在致命的侵襲性感染[5] 。煙曲霉具有完整的有性世代,可產生閉囊殼(cleistothecia)和子囊孢子。
隱球菌屬

新型隱球菌可以在艾滋病患者中引起嚴重形式的腦膜炎。大多數隱球菌屬都生活在土壤中,不會引起人類疾病。新型隱球菌是人類和動物的主要病原體。已知的羅倫隱球菌和淺白隱球菌偶爾會在免疫缺陷的人類患者中引起中度至重度疾病。格特隱球菌則是非洲大陸和澳大利亞的熱帶地區特有的,可以在免疫功能正常的人群中引起疾病[2]。
感染新型隱球菌細胞通常被肺內的肺泡巨噬細胞吞噬[6]。入侵的新型隱球菌細胞可能會被這些巨噬細胞釋放出氧化和亞硝化分子而被殺死[7]。然而,一些新型隱球菌細胞可能在巨噬細胞內存活[6]。病原體在巨噬細胞內存活的能力可能決定疾病的潛伏期,傳播和對抗真菌劑的抗性。為了在巨噬細胞惡劣的細胞內環境中存活,新型隱球菌的反應之一是增加用於響應氧化應激的基因表現[6]。
新型隱球菌的單倍體細胞核可以經歷細胞核融合以變成二倍體。然後這些二倍體核可能經歷減數分裂,包括重組,導致形成能夠分散的單倍體擔孢子[8]。減數分裂可能有助於修復新型隱球菌DNA以應對巨噬細胞的攻擊[8][9]。
組織胞漿菌
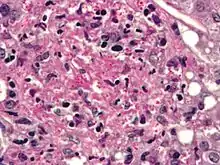
組織胞漿菌可導致人,狗和貓的組織胞漿菌病。這種真菌在美洲,印度和東南亞最為普遍,流行於美國的某些地區。感染通常是由於吸入受污染的空氣。
宿主防禦機制
參考資料
- . Nature Microbiology. 25 July 2017, 2 (8): 17120 [2017-11-11]. doi:10.1038/nmicrobiol.2017.120. (原始内容存档于2017-08-16) –nature.com.
- San-Blas G; Calderone RA (editors). . Caister Academic Press. 2008 [2014-12-29]. ISBN 978-1-904455-32-5 . (原始内容存档于2014-11-22).
- dEnfert C; Hube B (editors). . Caister Academic Press. 2007 [2010-05-28]. ISBN 978-1-904455-13-4 . (原始内容存档于2010-03-28).
- Machida, M; Gomi, K (editors). . Caister Academic Press. 2010. ISBN 978-1-904455-53-0.
- O'Gorman CM, Fuller H, Dyer PS. . Nature. 2009, 457 (7228): 471–4. PMID 19043401. doi:10.1038/nature07528.
- Fan W, Kraus PR, Boily MJ, Heitman J. . Eukaryotic Cell. 2005, 4 (8): 1420–33. PMC 1214536. PMID 16087747. doi:10.1128/EC.4.8.1420-1433.2005.
- Alspaugh JA, Granger DL. . Infect. Immun. 1991, 59 (7): 2291–6. PMC 258009. PMID 2050398.
- Lin X, Hull CM, Heitman J. . Nature. 2005, 434 (7036): 1017–21. PMID 15846346. doi:10.1038/nature03448.
- Bernstein H, Bernstein C, Michod RE (2018). Sex in microbial pathogens. Infection, Genetics and Evolution volume 57, pages 8-25. https://doi.org/10.1016/j.meegid.2017.10.024
- Ryan KJ; Ray CG (editors). 4th. McGraw Hill. 2004. ISBN 0-8385-8529-9.
- Robert, V. A.; Casadevall, A. . The Journal of Infectious Diseases. 2009, 200 (10): 1623–1626. PMID 19827944. doi:10.1086/644642.
- Brown, G. D., Drummond, R. A., Gaffen, S. L., & Hise, A. G. (2015). Innate Defense against Fungal Pathogens. Cold Spring Harb Perspect Med, 5(6). doi: 10.1101/cshperspect.a019620.
