苏门答腊犀
苏门答腊犀(学名:Dicerorhinus sumatrensis)简称 苏门犀,是 双角犀属 的唯一物种,现存体型最小的犀牛。肩高约 1.3 米,体长约 2.4-3.2 米,体重约 700 千克。苏门犀有两个角,与非洲的白犀和黑犀相似,而不同于亚洲的印度犀和爪哇犀。前角较大,通常为 15-25 厘米,後角较小,通常短于 10 厘米。身披厚密的红棕色长毛,是世界上最后生存的披毛犀牛。
| 蘇門答臘犀 | |
|---|---|
 | |
| 印尼楠榜省蘇門答臘犀保護區中的一头蘇門答臘犀 | |
| 保护状况 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 哺乳纲 Mammalia |
| 目: | 奇蹄目 Perissodactyla |
| 科: | 犀科 Rhinocerotidae |
| 亚科: | 犀亚科 Rhinocerotinae |
| 族: | 雙角犀族 Dicerorhinini |
| 属: | 雙角犀屬 Dicerorhinus Gloger, 1841 |
| 种: | 蘇門答臘犀 D. sumatrensis |
| 二名法 | |
| Dicerorhinus sumatrensis (Fischer, 1814)[3] | |
| 亞種 | |
D. s. lasiotis Buckland, 1872
D. s. sumatrensis Fischer, 1814 D. s. harrissoni Groves, 1965 | |
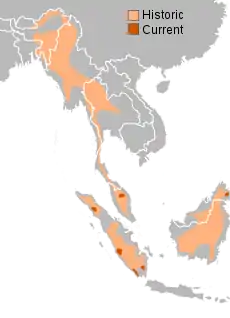 | |
| 蘇門答臘犀分布範圍 | |
苏门犀通常独居,仅在发情与扶养幼仔期间雌雄成对生活。它们是犀牛中发声最频繁的,通讯方式除了声音外,还会在泥中做记号或排便。
曾广泛分布于印度、不丹、孟加拉国、缅甸、老挝、泰国、马来西亚和印度尼西亚的热带雨林与沼泽中,历史上分布的最北端可至中国南部,尤其是四川[4][5] ,于 1916 年在中国灭绝。现仅存 4 个小而零散的野生种群(3 个在苏门答腊岛,1 个在婆罗洲北部,而马来半岛的种群已在近年灭绝),活体总数不到 80 头[6],是世界上濒危程度最高的动物之一。苏门犀濒危的主要原因是为其角而引发的偷猎。犀角在中药材中极其珍贵,每千克值 3 万美元。大規模伐木与森林的开发对苏门犀是巨大的威胁,且苏门犀在圈养环境下的适应性较差,人工繁殖的难度极高。
分类与命名
西方对苏门犀的最初记载是在1793年,当时一头犀牛在苏门答腊的Fort Marlborough十六公里外的郊野被枪杀。该标本的画像与描述记载被送到自然学家Joseph Banks的手里,当时英国皇家学会的总裁,他出版了一张关于此标本的文献。不过直到1814年这个物种才被德国科学家Johann Fischer von Waldheim正式命名。[7][8]
苏门犀的学名(Dicerorhinus sumatrensis)源于古希腊语的Di(两个), cero(角), rhinus (鼻), sumatrensis (苏门答腊的),其意思就是“苏门答腊的双鼻角犀”。[9]卡尔·林奈本将所有的犀牛分到犀牛属(Rhinoceros ),所以苏门犀最初的学名是(Rhinoceros sumatrensis)。后来的学家认为苏门犀的双角比较特别,并把它分到独立的属,双角犀属(Dicerorhinus)。
目前有三个亚种被认同:
- 苏门答腊亚种或西部亚种(D. s. sumatrensis)分布于苏门答腊岛与马来半岛。是现存数量最多的亚种。马来半岛的种群曾被列为独特的亚种D.s. niger,但之后发现它们与苏门答腊岛的犀牛十分相似。[3]
- 北方亚种(D. s. lasiotis)体型较大,曾广布于印度,印度支那与中国南部,现已灭绝。现在缅甸与云南仍有未证实的目击报告[10] 。近期在神农架发现石化程度很轻的骸骨,说明直到近代,这里依然有犀牛的存在。
近期基因研究发现了三个不同的种族。其中马来半岛与苏门答腊东部虽然被马六甲海峡分开,但它们的血缘关系较近。而苏门答腊西部的种群被巴里散山脉隔离的时间较长,婆罗洲的种群则更独特。亚种之间的交配不成问题。
进化
犀牛最早出现于早始新世。DNA的对比表明犀牛与马在5千万年前"分家"。[11][12]现存的犀科在晚始新世的欧亚大陆出现,现存犀牛的共同祖先生存在中新世。[13][14]
苏门犀是现存犀牛中最原始的一种,因为它保留了许多祖先的特征。最早的双角犀属的化石记录出现在中新世早期,大约2300-1600万年前。基因研究表明双角犀在2590±190万年前就已与其他犀牛分歧。目前有三种对于苏门犀与其他现存犀牛的亲缘关系的假说。一说苏门犀与非洲的黑犀与白犀是近亲,因为它们都有两只角。[11]其他分类学者认为苏门犀与印度犀和爪哇犀更接近,因为它们的分布区很接近。[11][15]近期研究则表明两种非洲犀牛,两种亚洲独角犀和苏门犀各自代表三种自2590万年前分裂的不同的世系,而哪一支先分歧目前仍不确定。[11][16]
由于形态的接近,苏门犀与著名的灭绝种类披毛犀(Coelodonta antiquitatis)被认为是近亲。近期的基因研究也支持这一说。披毛犀是著名的冰期动物,最早出现在中国。在晚更新世它们已广布于欧亚大陆,东起朝鲜西至西班牙,并于1万年前灭绝。[17]双角犀属的化石有很多,但苏门犀是唯一的近代物种。[18]
特征

一头典型的成年苏门犀肩高大约125厘米,体长在240-315厘米,平均体重大约700公斤,不过最大的个体可以达到1100公斤。苏门犀长有两个突角,这一点与非洲的白犀和黑犀相似,而不同于亚洲的印度犀和爪哇犀。前角较大,通常为15-25厘米,不过最大的记录据说有81厘米,后角较小,通常是少于10厘米。[18]角的颜色呈黑灰色或黑色,雄性的角大于雌性,不过此外两性没有区别。野外寿命大约为30-45年,饲养环境中的记录是一头雌性的北方苏门犀,活了32年与8个月并在1900年死于伦敦动物园。
四腿之后有两片褶皱的皮肤,颈部有一片较小的皱肤。皮肤较薄,有2-3厘米厚。野外个体没有皮下脂肪。毛有浓有稀,幼仔的毛较密,毛色为红棕色。野外的毛较难观察因为它们身上通常覆盖着泥。饲养个体的毛较濃密因为与草木的摩擦较少。耳中的毛较长,尾尖有一簇浓密的毛。与其他犀牛一样,苏门犀的视力很差。虽然它们看似笨重,但其實它们可以迅速的在叢林中穿梭。[19][18][20]
栖息与分布

苏门犀在低地与高原都有出现,生活在雨林,沼泽和雨林中。栖息于接近水源的丘陵地带,尤其是灌木较浓的山坡地带。苏门犀曾经遍布于苏门答腊,婆罗洲,印度,印度支那和中国华南甚至中原地区(中国早期的记载并没有鉴别种类,所以并不确定是否苏门犀和爪哇犀两种都有。从现有文物来看,如小臣艅犀尊 、战国错金银云纹铜犀尊均为双角,似以苏门犀为主)。
2019年5月27日,大馬最後一隻雄蘇門犀「塔姆」(Tam)離世[21],同年11月23日,大馬最後一隻雌蘇門犀「伊曼」(Iman)離世。至此苏门答腊犀牛仅在苏门答腊与婆罗洲有零星分布。[22]未证实的记载在各地都有,不过缅甸有存活种群的证据,可惜该国的政治混乱阻止了进一步的调查。[23]
习性
苏门犀是独居动物,雄性的生活范围可达50平方公里,雌性为10-15平方公里。雌雄的地盘经常重叠。目前没有发现因争夺地盘而搏斗的行为。它们会挖泥土,排便,或把树苗弯成独特的样式来表示它们的地盘。通常黄昏和清晨出来觅食,白天待在泥池里休息。在雨季会迁徙到海拔较高的地区,在较寒冷的季节迁回山谷[19]。

苏门犀花很长一段时间泡泥澡。当泥坑变小时,它们会用角与脚去挖深。泥坑会被用上2-12周,然后被抛弃。它们通常在中午泡上2-3个小时。泥澡能帮助它们维持体温并去除皮外寄生虫。苏门犀的人工饲养之所以比较容易失败,正是因为泥澡的缺乏,导致皮肤发炎与化脓,眼病,掉毛以及最终的死亡。[24]
饲养的苏门犀的主要死于虱子与胃蝇[20]。它们也会受苏拉病的影响,寄生锥体虫受到苍蝇的传播。在2004年的18天内,有5头犀牛得此病而陆续死亡。除了人类之外,成年个体没有天敌。幼仔会受到虎、豺和野狗的攻击。虽然苏门犀与亚洲象和马来貘的分布重叠,它们之间并没有食物竞争。它们甚至会公用一条道路。[19][25]
食性
 |
 |
 |
 |
| 苏门犀的食性很广,包括(从左上角顺时针):野桐属、山竹、紫金牛属和番樱桃属[25][26]。 | |
通常在黄昏和清晨出来觅食。以小嫩叶、树苗、水果、嫩芽和细枝为食。[18]每天可吃掉50公斤。[19]从粪便标本可看出,苏门犀的以超过100种植物为食。最常见的食物是树干直径1-6厘米的树苗。它们通常将这些树苗推倒,并吃掉上面的树叶。最常见的被食种类为番樱桃属的植物。
苏门犀的食物含大量的膳食纤维与适中的蛋白质。[27]盐是很重要的营养来源,进行盐田的地方包括小热泉,咸水的渗漏或泥热泉。盐田也存在着社交目的,雄性会巡视盐田来寻找雌性。有些犀牛生存的地区不存在盐田,它们靠吃些含许多矿物的植物来获得必需的营养。[25][26]
交流
| Sumatran Rhinoceros vocalizations (.wav files)[28] | |
|---|---|
苏门犀是犀牛中最健谈的,[28]饲养和野外的观察发现它们几乎不停的发声。[20] 最常见的三种声音包括“咦”,鲸歌与尖哨声。“咦”是一种长约1秒的尖叫,是最常见的声音。它们也会发出与露脊鲸相似的声音,故名鲸歌。尖哨声是三种声音中最响的,响到能让动物园的铁栏震动。这些声音的目的目前还不清楚,不过应该与其他有蹄类一样,用来警报、表达位置和性问题。尖哨声可以在很远的距离被听见。亚洲象发出的响声相近地声音可散播至9.8km以外,因此苏门犀的尖哨声应该也能传播至相似的距离。[28]苏门犀会将没有吃掉的树苗扭曲,以表明自己的地盘或小径的交叉口。[25]
繁殖
雌性在6-7岁达到性成熟,雄性则要达到10岁以后。孕期为15-16个月。幼仔体重通常为40-60公斤,在15个月后断奶,并在1-3岁时待在母亲身边。在野外生殖间歇期为4-5年。目前我们对幼仔抚养的行为并不了解。
苏门犀的性习性在饲养环境中被了解。性关系之前会出现一段恋爱期,导致声音交流的增加,尾部的提高,小便以及更多的肉体接触,包括雌雄体用口鼻部抚摸互相的头部与生殖器。恋爱的习性与黑犀较相似。年轻的雄犀牛往往会对雌性施暴,导致雌性的受伤甚至死亡。在野外雌性会逃离过度粗暴的雄性,在饲养的笼圈中则不行,导致较低的饲养生殖率。[29][30][31]
发情期长约24小时,并会在21-25天后复发。辛辛纳提动物园的个体交配时间大约为30-50分钟,与其他犀牛相似,在马来西亚的苏门犀保护中心的个体则较短暂。苏门犀较长的交配时间与对其他犀牛的观察,表明其交配季节应该也比较长。[29]2001年之前的饲养繁殖都因各种原因而失败。研究发现苏门犀的排卵靠交配的催生且有不稳定的孕酮数量。[32]2001年的成功是因为孕酮的补给。[33]
濒危与保护
苏门犀在东南亚的数量曾经很多,现在野外不足百头。活体总数可能比爪哇犀略多,但苏门犀灭绝的威胁更大,因为当地较严重的捕猎与栖息地的丧失,而且它们的种群相隔比较远。相反几乎所有的爪哇犀都生活在爪哇岛的Ujung Kulon半岛。另外苏门犀的数量正在下降,而爪哇犀的数量则比较稳定。两种犀牛都被列为极危动物。偷猎与栖息地的丧失导致它们仅存于人类几乎无法到达的地域。[34][35]
虽然针对苏门犀的捕猎没有非洲犀牛那么严重(至少在捕猎的数量上),但它依然是个问题。犀牛角的价格极其昂贵,可达每公斤3万美元,且价格会随着数量的减少而提高。[13]苏门犀并不像非洲犀样受到大规模的捕杀,因为它们很难被直接捕猎或研究(一名研究者在一盐田蹲了7个星期才遇到活的犀牛)。陷阱的使用则更常见。当地人用犀角当护身符,认为它可防止疾病与中毒。干犀肉则被用来当治腹泻,麻风与肺结核的药材。“犀油”会在犀牛头骨在椰油泡上数星期后生产,并用来治皮肤病。这些材料在中医药学中使用甚广。[20][23][25] 犀牛部位曾被当做春药使用。[13]
印度尼西亚与马来西亚的热带雨林是多数合法与违法的伐木目标,由于其硬木树制的可贵性。稀有树种像印茄木,梅兰蒂木与胶木是珍贵的国际市场产品,其价格可达每立方米1800美元。非法伐木业的组织很难进行,因为许多人类生活与犀牛同样的森林。
參考文献
- van Strien, N.J., Manullang, B., Sectionov, Isnan, W., Khan, M.K.M, Sumardja, E., Ellis, S., Han, K.H., Boeadi, Payne, J. & Bradley Martin, E. 2008. 页面存档备份,存于 Dicerorhinus sumatrensis. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.2.
- Dicerorhinus sumatrensis Convention on International Trade in Endangered Species
- Rookmaaker, L.C. . Journal of the Malayan Branch of the Royal Asiatic Society. 1984, 57 (1): 12–25.
- The Art of Rhinoceros Horn Carving in China (1999), p. 27. Jan Chapman. Christie’s Books, London.
- The Golden Peaches of Samarkand: A study of T’ang Exotics (1963), p 83. Edward H. Schafer. University of California Press. Berkeley and Los Angeles. First paperback edition: 1985.
- . 蘋果日報. 2019-05-28.
- Rookmaaker, Kees. . Fulconis, R. (编). . London: European Association of Zoos and Aquaria. 2005: 52.
- Morales, Juan Carlos; Patrick Mahedi Andau, Jatna Supriatna, Zainuddin Zainal-Zahari, and Don J. Melnick. . Conservation Biology. 1997, 11 (2): 539–543. doi:10.1046/j.1523-1739.1997.96171.x.
- Liddell, Henry G.; and Robert Scott. Abridged. Oxford: Oxford University Press. 1980. ISBN 0-19-910207-4.
- Asian Rhino Specialist Group (1996). Dicerorhinus sumatrensis ssp. lasiotis 页面存档备份,存于. 2007 IUCN Red List of Threatened Species. IUCN 2007. Retrieved on January 13, 2008.
- Tougard, C.; T. Delefosse, C. Hoenni, and C. Montgelard. . Molecular Phylogenetics and Evolution. 2001, 19 (1): 34–44. doi:10.1006/mpev.2000.0903.
- Xu, Xiufeng; Axel Janke, and Ulfur Arnason. . Molecular Biology and Evolution. 1 November 1996, 13 (9): 1167–1173 [2007-11-04]. (原始内容存档于2010-08-08).
- Dinerstein, Eric. . New York: Columbia University Press. 2003. ISBN 0-231-08450-1.
- Lacombat, Frédéric. . Fulconis, R. (编). . London: European Association of Zoos and Aquaria. 2005: 46–49.
- Groves, C. P. . Zeitschrift fuer Zoologische Systematik und Evolutionsforschung. 1983, 21: 293–313.
- Cerdeño, Esperanza, (PDF), Novitates (American Museum of Natural History), 1995, (3143) [2010-01-29], ISSN 0003-0082, (原始内容存档 (PDF)于2009-03-27)
- Orlando, Ludovic; Jennifer A. Leonard, Aurélie Thenot, Vincent Laudet, Claude Guerin, and Catherine Hänni. . Molecular Phylogenetics and Evolution. September 2003, 28 (2): 485–499. doi:10.1016/S1055-7903(03)00023-X.
- Groves, Colin P., and Fred Kurt. (PDF). Mammalian Species. 1972, (21): 1–6 [2010-01-29]. doi:10.2307/3503818. (原始内容存档 (PDF)于2016-03-03).
- van Strien, Nico. . Fulconis, R. (编). . London: European Association of Zoos and Aquaria. 2005: 70–74.
- van Strien, N.J. . Mededelingen Landbouwhogeschool Wageningen. 1974, 74 (16): 1–82.
- . 星洲网. 2019-05-28.
- . BBC. 2019-11-23 (英语).
- Foose, Thomas J. and van Strien, Nico, , IUCN, Gland, Switzerland, and Cambridge, UK, 1997, ISBN 2-8317-0336-0
- Julia Ng, S.C.; Z. Zainal-Zahari, and Adam Nordin. . Journal of Wildlife and Parks. 2001, 19: 7–12.
- Borner, Markus. . Zurich : Juris Druck & Verlag. 1979. ISBN 3260046003.
- Lee, Yook Heng; Robert B. Stuebing, and Abdul Hamid Ahmad. . Biotropica. 1993, 3 (5): 352–355. doi:10.2307/2388795.
- Dierenfeld, E.S.; A. Kilbourn, W. Karesh, E. Bosi, M. Andau, S. Alsisto. . Zoo Biology. 2006, 25 (5): 417–431. doi:10.1002/zoo.20107.
- von Muggenthaler, Elizabeth; Paul Reinhart, Brad Limpany, and R. Barton Craft. . Acoustics Research Letters Online. 2003, 4 (3): 83. doi:10.1121/1.1588271.
- Zainal Zahari, Z.; Y. Rosnina, H. Wahid, K.c. Yap, and M.R. Jainudeen. . Animal Reproduction Science. 2005, 85: 327–335. doi:10.1016/j.anireprosci.2004.04.041.
- Zainal-Zahari, Z.; Y. Rosnina, H. Wahid, and M. R. Jainudeen. . Anatomia, Histologia, Embryologia: Journal of Veterinary Medicine Series C. 2002, 31 (6): 350–354. doi:10.1046/j.1439-0264.2002.00416.x.
- Roth, T.L.; R.W. Radcliffem, and N.J. van Strien. . International Zoo News. 2006, 53 (6): 352–353.
- Roth, T.L.; J.K. O'Brien, M.A. McRae, A.C. Bellem, S.J. Romo, J.L. Kroll and J.L. Brown. . Reproduction. 2001, 121: 139–149. PMID 11226037. doi:10.1530/rep.0.1210139.
- Roth, T.L. . Hormones and Behavior. 2003, 44: 31.
- Rabinowitz, Alan. . Conservation Biology. 1995, 9 (3): 482–488. doi:10.1046/j.1523-1739.1995.09030482.x.
- van Strien, Nico J. . Proceedings of the International Elephant and Rhino Research Symposium, Vienna, June 7–11, 2001 (Scientific Progress Reports). 2001.
{kind=link}