11β-羟基类固醇脱氢酶
11β-羟基类固醇脱氢酶(HSD-11β或11β-HSD)是一类催化惰性十一酮产物(可的松)转化为活性的皮质醇或其逆反应的酶,从而调节糖皮质激素进入类固醇受体的途径:
- 11β-羟基类固醇 + NADP+ ⇌ 11-甾酮 + NADPH + H+
| 11β-羟基类固醇脱氢酶(NADP+) | |||||||
|---|---|---|---|---|---|---|---|
 | |||||||
| 11β-羟基类固醇脱氢酶1,二聚体,人 | |||||||
| |||||||
| 识别码 | |||||||
| EC编号 | 1.1.1.146 | ||||||
| CAS号 | 9041-46-7 | ||||||
| 数据库 | |||||||
| IntEnz | IntEnz浏览 | ||||||
| BRENDA | BRENDA入口 | ||||||
| ExPASy | NiceZyme浏览 | ||||||
| KEGG | KEGG入口 | ||||||
| MetaCyc | 代谢路径 | ||||||
| PRIAM | 概述 | ||||||
| PDB | RCSB PDB PDBj PDBe PDBsum | ||||||
| 基因本体 | AmiGO / EGO | ||||||
| |||||||
因此,这种酶的两种底物是11β-羟甾体和NADP+, 而它的3种产物是11-甾酮、NADPH和H+。
这种酶属于氧化还原酶家族,特别是那些以NAD+或NADP+为受体的以CH-OH基团为供体的酶。该酶类的系统名称是11-β羟基类固醇:NADP+ 11-氧化还原酶。其他常用名称包括皮质类固醇11β-脱氢酶、β-羟基类固醇脱氢酶、11β-羟基类固醇脱氢酶、皮质类固醇11-还原酶, 和脱氢酶,11β-羟基类固醇。该酶参与c21-甾体激素代谢及雄激素和雌激素代谢。
功能
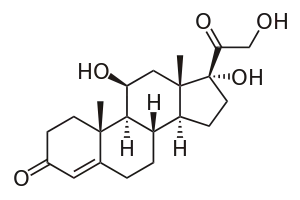
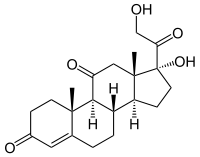
皮质醇,一种糖皮质激素,与糖皮质激素受体结合。 然而,由于其与醛固酮的分子相似性,它也在较高浓度下与盐皮质激素受体结合。醛固酮和皮质醇对盐皮质激素受体有相似的亲和力;然而,循环中的皮质醇远远多于醛固酮。为了防止皮质醇对盐皮质激素受体的过度刺激,11β-HSD将具有生物活性的皮质醇转化为不再与盐皮质激素受体结合的非活性皮质醇。11β-HSD与肾上腺皮质激素受体共定位。甘草含有甘草酸和甘草次酸,能抑制11β-HSD,导致盐皮质激素过多综合征。皮质醇水平随之升高,皮质醇与盐皮质激素受体结合产生低钾血症、碱中毒和高血压(即盐皮质激素过量)的临床症状和体征。
亚型
| HSD11B1 | 依赖NADPH | 在包括肝脏、脂肪組織和中枢神经系统的关键代谢组织中高度表达。 | 在这些组织中,HSD11B1将可的松还原为能激活糖皮质激素受体的活性激素皮质醇。 |
| HSD11B2 | 依赖NAD+ | 在醛固酮选择性组织中表达,包括肾脏、肝脏、肺、结肠、唾液腺、HSD2神经元和胎盘。 | 在这些组织中,HSD11B2将皮质醇氧化为可的松,并防止盐皮质激素受体的异常激活。 |
临床应用
11β-羟基类固醇脱氢酶(HSD11B)是一种参与类固醇激素生理过程的酶。11β-羟基类固醇脱氢酶存在两种亚型,即HSD11B-1型和HSD11B-2型。在糖皮质激素靶向的代谢组织中发现1型亚型,其将皮质醇转化为活性皮质醇。[3] HSD11B-1型可作为一种还原酶,产生活性皮质醇和放大糖皮质激素的作用。这种酶在肝脏中最丰富,但在身体的大多数组织中都能找到。HSD11B的增加是内脏肥胖的常见机制。[4]HSD11B-1可增强肝脏和脂肪组织中的糖皮质激素浓度,糖皮质激素过量可导致肥胖,并伴有高血压和糖尿病等其他特征。[5] 2型亚型由醛固酮选择性组织表达,并通过使用酶11-氧还原酶将其转化为皮质醇来保护盐皮质激素受体免受皮质醇激活。2型通过降低局部皮质醇水平和防止11-氧还原酶激活来保护组织免受持续激活。[3] 在不表达盐皮质激素受体的组织中,如胎盘和睾丸,它保护细胞免受皮质醇的生长抑制和/或促凋亡作用,特别是在胚胎发育期间。该基因突变引起明显的盐皮质激素过多和高血压综合征。[6]
对11β脱氢酶进行了研究,发现它能影响多种组织和机体功能。由于这种酶的主要功能是调节糖皮质激素,它与各种过度刺激或消耗糖皮质激素有关,从而导致人体内的化学失衡。11β脱氢酶的临床应用在肥胖相关研究和妊娠研究方面取得了重大进展。酶的作用、涉及到的特定的身体功能和相关的疾病如下。
高脂血症对11β-羟类固醇脱氢酶的影响
高脂血症对11β-羟类固醇脱氢酶有很大影响。[7]糖皮质激素依赖于血浆糖皮质激素浓度、细胞糖皮质激素受体表达和11β-HSD催化的受体前激素代谢。[7]控制皮质醇浓度的11β-羟基类固醇脱氢酶有两种:11β-HSD1和11β-HSD2。[7] 11β-HSD1作为氧还原酶负责将可的松转化为皮质醇,因为它依赖于NADP(H),而11β-HSD2通过NAD使皮质醇失活为可的松。[7]10-d高脂血症增加内脏和皮下脂肪组织中11β-HSD1的表达。[7]高脂血症降低肝脏和脂肪组织中11β-HSD2的表达。[7]高脂血症对11β-HSD1和11β-HSD2有重要影响。[7] 这说明高脂血症可能与皮质醇代谢有关。[7]
11β-HSD在器官中的活性
11β-羟基类固醇脱氢酶在器官和肾上腺中都有活性。[8]这种酶的亚型承担着不同的职责。[8]在活跃状态下,第一种亚型促进肝细胞内糖皮质激素的增加,并促进糖异生。[8] 第二种亚型将活性糖皮质激素转化为靶组织(如肾脏、唾液腺、肠道等)中的非活性代谢产物。[8]肾脏和肝脏中11β-HSD的两种亚型的激活触发了肾上腺外四氧嘧啶糖尿病的形成, 与肾上腺糖皮质激素合成减少有关。[8]肾上腺外的形成导致肝内皮质酮的局部形成增加,并伴随糖异生反应活跃。[8]这些糖异生反应增强了类似于糖尿病的持续代谢紊乱。[8] 因为其使局部皮质酮的增加,11β-HSD的第一个亚型可作为糖尿病、肥胖症和代谢综合征的潜在治疗药物。[8]
大脑中的作用
11β-羟基类固醇脱氢酶在老年人中枢神经系统中表达。[9]它在下丘脑-垂体-肾上腺轴功能中是必不可少的。[9] 11β-羟甾体脱氢酶也参与了衰老引起的意识智力活动下降。[9]这种酶也在发育阶段有助于中枢效应。[9] 例如,11β-羟基类固醇脱氢酶2型经常出现在胎儿组织中,如新生儿的大脑和胎盘。[9]如果11β-HSD2在胎儿组织中存在缺失或下降,则存在焦虑等不利发展后果。[9]
11β-羟基类固醇脱氢酶部分负责细胞内代谢,决定细胞内糖皮质激素的运作。[9]糖皮质激素影响大脑发育,最终影响中枢神经系统的功能。[9]如此重要,以至于一旦过剩或数量不足,后果是畸形贯穿一生。[9]11β-HSD1负责激活糖皮质激素,11β-HSD2负责使糖皮质激素失活。[9]11β-HSD1激活糖皮质激素的后果是认知能力下降,特别是随着年龄的增长。[9] 相反,11β-HSD2的作用发生在发育过程中。[9]高表达11β-HSD2的一些后果是焦虑和心脏代谢紊乱,两者都是早期糖皮质激素程序设计的一部分。[9]
参见
- 类固醇生成酶
- 11β-羟基类固醇脱氢酶1型
- 11β-羟基类固醇脱氢酶2型
- 可的松还原酶缺乏症
参考文献
- Seckl JR. . Front Neuroendocrinol. January 1997, 18 (1): 49–99. PMID 9000459. doi:10.1006/frne.1996.0143.
- Anagnostis P, Athyros VG, Tziomalos K, Karagiannis A, Mikhailidis DP. . The Journal of Clinical Endocrinology and Metabolism. 2009, 94 (8): 2692–2701 [2011-03-13]. PMID 19470627. doi:10.1210/jc.2009-0370. (原始内容存档于2010-07-12).
- Lindsay, Kaitlin. . kaitlinlindsay.com. (原始内容存档于2019-04-22).
- Ellis, Wendy L. (PDF). semanticscholar.org. [November 23, 2018]. (原始内容存档 (PDF)于2019-03-03).
- Hughes, Katherine A; Webster, Scott P; Walker, Brian R. . Expert Opinion on Investigational Drugs. 2008-03-25, 17 (4): 481–496. ISSN 1354-3784. PMID 18363514. doi:10.1517/13543784.17.4.481 (英语).
- . www.greencards.org. (原始内容存档于2011-06-02).
- Sieber-Ruckstuhl, Nadja S.; Zini, Eric; Osto, Melanie; Franchini, Marco; Boretti, Felicitas S.; Meli, Marina L.; Sigrist, Brigitte; Lutz, Thomas A.; Reusch, Claudia E. (PDF). Domestic Animal Endocrinology. November 2010, 39 (4): 222–230 [2020-01-28]. ISSN 0739-7240. PMID 20688460. doi:10.1016/j.domaniend.2010.06.003. (原始内容存档 (PDF)于2018-07-19).
- Cherkasova, O. P.; Selyatitskaya, V. G.; Pal’chikova, N. A.; Kuznetsova, N. V. . Bulletin of Experimental Biology and Medicine. 2014-11-29, 158 (2): 185–187. ISSN 0007-4888. PMID 25430643. doi:10.1007/s10517-014-2718-3 (英语).
- Wyrwoll, Caitlin S.; Holmes, Megan C.; Seckl, Jonathan R. . Frontiers in Neuroendocrinology. August 2011, 32 (3): 265–286. ISSN 0091-3022. PMC 3149101. PMID 21144857. doi:10.1016/j.yfrne.2010.12.001.
- Rogers, Samantha L.; Hughes, Beverly A.; Jones, Christopher A.; Freedman, Lauren; Smart, Katherine; Taylor, Norman; Stewart, Paul M.; Shackleton, Cedric H. L.; Krone, Nils P. . The Journal of Clinical Endocrinology & Metabolism. May 2014, 99 (5): E821–E831. ISSN 0021-972X. PMID 24517145. doi:10.1210/jc.2013-3254 (英语).
- Agarwal AK, Monder C, Eckstein B, White PC. . J. Biol. Chem. 1989, 264 (32): 18939–43. PMID 2808402.
- Bush IE, Hunter SA, Meigs RA. . Biochem. J. 1968, 107 (2): 239–58. PMC 1198650. PMID 4384445. doi:10.1042/bj1070239.
- Lakshmi V, Monder C. . Endocrinology. 1988, 123 (5): 2390–8. PMID 3139396. doi:10.1210/endo-123-5-2390.
- Phillips DM, Lakshmi V, Monder C. . Endocrinology. 1989, 125 (1): 209–16. PMID 2661206. doi:10.1210/endo-125-1-209.