血清素
血清素(英語:,全稱血清張力素,又稱5-羟色胺和血清胺,简称为5-HT)為單胺型神經遞質,由色氨酸经色氨酸羟化酶转化为5-羟色氨酸,再经5-羟色氨酸脱羧酶在中樞神經元及動物(包含人類)消化道之腸嗜鉻細胞中合成。5-羥色胺主要存在於動物(包括人類)的胃腸道、血小板和中樞神經系統中。它被普遍認為是幸福和快樂感覺的貢獻者。血清素在大脑中的含量为总量的2%,有九成位于粘膜肠嗜鉻细胞和肌间神经丛,参与肠蠕动的调节[4][5]。与肠粘膜进入血液的5-HT主要被血小板摄取。8%-9%的位于血小板中。因为5-HT不能通过血脑屏障,故中枢和外周可视为两个独立的系统。
| 血清素(Serotonin) | |
|---|---|
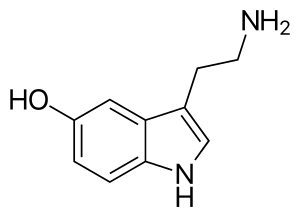 | |
| IUPAC名 | 3-(2-Aminoethyl)indol-5-ol |
| 同义词 | 5-羟色胺,5-Hydroxytryptamine, Enteramine; Thrombocytin, 3-(β-Aminoethyl)-5-hydroxyindole, Thrombotonin |
| 缩写 | 5-HT |
| 源 | 中缝核, 肠嗜铬细胞 |
| 靶 | 广泛 |
| 受体 | 5-HT1, 5-HT2 受体, 5-HT3, 5-HT4 受体, 5-HT5, 5-HT6, 5-HT7 |
| 激动剂 | SSRI, MAOI (间接) |
| 前体 | 5-HTP |
| 合成酶 | 芳族L-氨基酸脱羧酶 |
| 代谢酶 | 单胺氧化酶 |
| 数据库链接 | |
| CAS注册号 | 50-67-9 |
| PubChem | CID: 5202 |
| ChemSpider | 5013 |
| KEGG | C00780 |
| 血清素 | |
|---|---|
 | |
 | |
| IUPAC名 5-Hydroxytryptamine or 3-(2-Aminoethyl)indol-5-ol | |
| 别名 | 5-Hydroxytryptamine, 5-HT, Enteramine; Thrombocytin, 3-(β-Aminoethyl)-5-hydroxyindole, Thrombotonin |
| 识别 | |
| CAS号 | 50-67-9 |
| PubChem | 5202 |
| ChemSpider | 5013 |
| SMILES |
|
| InChI |
|
| InChIKey | QZAYGJVTTNCVMB-UHFFFAOYAX |
| ChEBI | 28790 |
| KEGG | C00780 |
| MeSH | Serotonin |
| IUPHAR配体 | 5 |
| 性质 | |
| 化学式 | C10H12N2O |
| 摩尔质量 | 176.215 g/mol g·mol⁻¹ |
| 外观 | White powder |
| 熔点 | 167.7 °C(441 K) |
| 沸点 | 416 ± 30 °C |
| 溶解性(水) | slightly soluble |
| pKa | 10.16 in water at 23.5 °C[1] |
| 偶极矩 | 2.98 D |
| 危险性 | |
| 致死量或浓度: | |
LD50(中位剂量) |
750 mg/kg (subcutaneous, rat),[2] 4500 mg/kg (intraperitoneal, rat),[3] 60 mg/kg (oral, rat) |
| 若非注明,所有数据均出自一般条件(25 ℃,100 kPa)下。 | |
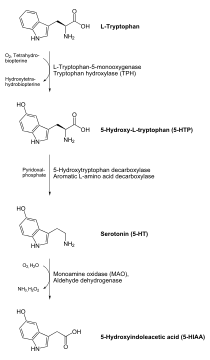
人體大約90%的總5-羥色胺位於腸胃道中的嗜鉻細胞中,它用於調節腸的蠕動。5-羥色胺分泌於腸管和基底面,由此增加了血小板對血清素的吸收。5-羥色胺激活後增加刺激 myenteric plexus影響腸蠕動的速率。剩餘部分在中樞神經的血清素能神經元中合成,其中它具有各種功能,這些包括調節心情,食慾和睡眠。血清素還具有一些認知功能,包括記憶和學習。在突觸處調節5-羥色胺,被認為是幾類抗抑鬱藥藥物的主要作用。
嗜鉻細胞分泌的血清素最終從組織中出來進入血液中。它由血小板積極吸收與存儲它。當血小板凝結成塊時,血小板釋放血清素,其用作血管收縮劑並有助於調節血液凝固和止血。血清素也是某些細胞的生長因子,其在傷口癒合中起到作用。有各种血清素受體。
5-羥色胺主要由肝臟代謝為5-羥基吲哚乙酸(5-HIAA)。代謝包括首先通過單胺氧化酶氧化成相應的醛。然後通過醛脫氫酶氧化成5-羥基吲哚乙酸(5-HIAA),一種吲哚乙酸衍生物。然後後者由腎臟排出。
除了動物,在真菌和植物中也發現5-羥色胺。5-羥色胺在昆蟲毒液和植物的刺中存在,會引起疼痛,這也是注射5-羥色胺時的副作用。血清素由致病性變形蟲產生,其對腸道的影響是引起腹瀉。其廣泛存在於許多種子和果實中可能有助於刺激消化道排出種子,幫助植物播種。
許多真菌與植物中皆含有血清素[6],而人类必须通过食物获取色氨酸。
5-HT的影响涉及多个生理系统,特别与觉醒水平(中國用語),睡眠-觉醒周期,心境食物和性行为有密切相关。有动物实验表明,当提高血清素在动物体内含量时,动物的互相攻击行为明显减少。
血清素是一种抑制性神经递质,最早于血清中发现,广泛存在于哺乳动物组织中,在大脑皮层质及神经突触内含量很高。在外周组织,血清素是一种强血管收缩剂和平滑肌收缩刺激剂。血清素还能增强记忆力,并能保护神经元免受“兴奋神经毒素”的损害。如谷氨酸即对受损的神经细胞有很大的毒性,因此充足的血清素能在老化过程中防止脑损害发生。
作用机制
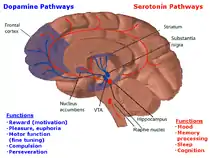
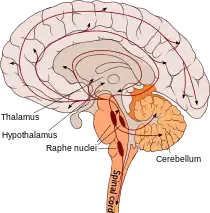
脑干中的中缝核是哺乳动物脑5-HT神经元密度最大的核团,大脑皮层是主要的投射区域,其中投射到额叶皮层的神经纤维最为浓密。还有枪额叶皮层以及运动皮层,主要投射来自于背侧中缝核。正中缝合和背侧中缝核的5-HT神经元包含高度并行的神经纤维,投射到多个终端领域。这表明那些在功能上相关的核可能接受同一组5-HT神经元甚至同一个神经元的投射。5-HT受体的现代分类是基于它的结构特征和使用的第二信使系统。5-HT必须通过相应的受体的介导方能产生作用。5-HT受体复杂,已发现7种5-HT受体亚型。其中仅5-HT3受体与配体门控通道离子通道偶联,其余6种均与G蛋白偶联,他们的结构包括7个跨膜区段,3个胞浆环和3个细胞外环。5-H通过激动不同的5-HT受体,可具有不同药理作用。
- 心血管系统:作用复杂,静脉注射微克5-HT可引起血压的三相反应:
- 短暂的降低,这与5-HT激动5-HT3受体,引起心脏负性频率作用有关;
- 持续数分钟高血压,这是5-HT激动5-HT2受体,引起肾、肺等组织血管收缩所致;
- 长时间的低血压,是骨骼肌血管舒张所致,需要血管内皮细胞的参与。此外,5-HT激动血小板5-HT2受体,可引起血小板聚集。
- 平滑肌:5-HT激动胃肠道平滑肌5-HT2受体,或肠壁内神经节细胞5-HT4受体,均可以引起胃肠道平滑肌收缩,使胃肠道张力增加,肠蠕动加快;5-HT尚可兴奋支气管平滑肌,哮喘病人对其特别敏感,但对正常人影响较小。
- 神经系统:动物侧脑室注射5-HT后,可引起镇静、嗜睡和一系列行为反应,并影响体温调节和运动功能。虫咬和某些植物的刺颗刺激5-HT释放,作用于感觉神经末梢,引起痒、痛。5-HT本身尚无临床应用价值。临床上使用选择性5-羟色胺再吸收抑制剂(SSRIs)和单胺氧化酶抑制剂(MAOIs)提高突触间隙5-HT浓度以治疗抑郁强迫诸多精神症状。但存在着引发血清素综合症的风险。
细胞作用
受体
血清素受体:或稱5-羥色胺受體位于动物神经细胞和其它类型细胞的细胞膜,並介導血清素作為內源性配體和廣泛範圍的藥物和致幻藥物的作用。除了5-HT3受體,配體門控離子通道(LGIC),所有其他血清素受體是G蛋白偶聯受體(GPCR),其激活細胞內第二信使級聯。(也稱為七跨膜受體或七螺旋受體)。
血清素受体是在中樞和周圍神經系統中發現的一組G蛋白偶聯受體(GPCR)和配體門控離子通道(LGIC)。他們同時介導「興奮性」和「抑制性」神經傳遞。血清素受體被激活神經遞質「血清素」,其充當它們的天然配體。
血清素受體調節許多神經遞質,包括谷氨酸,γ-胺基丁酸(GABA),多巴胺,腎上腺素/去甲腎上腺素和乙酰膽鹼,以及許多其他激素,包括催產素,催乳素,加壓素,皮質醇,促腎上腺皮質激素和 P物質等等。血清素受體影響各種生物學和神經學過程,例如攻擊,焦慮,食慾,認知,學習,記憶,情緒,噁心,睡眠和體溫調節。血清素受體是多種藥物的靶標,包括許多抗抑鬱藥,抗精神病藥,減食慾藥,止吐藥,胃胃動力藥,抗偏頭痛藥,致幻劑和放心藥。
血清素受體幾乎在所有的動物和人中都可以發現,甚至已知在原始線蟲、秀麗隱桿線蟲(Caenorhabditis elegans),調節長壽和行為老化。
科
血清素受體可分為七個亞科 5-HT1, 5-HT2, 5-HT3, 5-HT4, 5-HT5, 5-HT6, 5-HT7。至少有十四種受體亞型已被發現,包含G蛋白偶聯受體和配體門控離子通道(G protein-coupled receptor and a ligand-gated ion channe)。5-HT1受體是5-HT受體家族中最龐大的一科,目前有5種亞型:A、B、D、E、F。5-HT2受體有A、B、C, 3種亞型。
| 家族 | 類型 | 作用機制 | 類別 |
| 5-HT1 | Gi/Go-蛋白偶聯受体 | 減少cAMP在細胞內的水平 | 抑制性 |
| 5-HT2 | Gq/G11-蛋白偶聯受体 | 增加細胞內IP3和DAG的水平 | 興奮性 |
| 5-HT3 | 配體門控Na+,K+离子通道 | 使質膜去極化 | 興奮性 |
| 5-HT4 | Gs蛋白偶聯受体 | 增加細胞內cAMP的水平 | 興奮性 |
| 5-HT5 | Gi/Go-蛋白偶聯受体[7] | 減少cAMP在細胞內的水平 | 抑制性 |
| 5-HT6 | Gs-蛋白偶聯受体 | 增加細胞內cAMP的水平 | 興奮性 |
| 5-HT7 | Gs-蛋白偶聯受体 | 增加細胞內cAMP的水平 | 興奮性 |
亞型
- 5-HT1受體是5-HT受體家族中最龐大的一科,目前有A、B、D、E、F5種亞型, 5-HT1A、5-HT1B、5-HT1D、5-HT1E、5-HT1F五種受體蛋白。
- 5-HT2受體次家族有A、B、C, 3種亞型, 5-HT2A、5-HT2B,和5-HT2C三種受體蛋白。
- 5-HT3受體
- 5-HT4受體
- 5-HT5受體次家族有A、B、2種亞型, 5-HT5A、5-HT5B二種受體蛋白。
- 5-HT6受體
- 5-HT7受體
總共有十四種受體亞型。
5-HT1A 受體
5-HT1A 受體是所有5-HT受體中最廣泛的。 在中樞神經系統中,5-HT1A受體以高密度存在於大腦皮層,海馬,腦隔膜,杏仁核和中縫核中,也以低量存在於基底神經節和丘腦中。
神經調節功能 5-HT1A 受體激動劑參與神經調節。 它們通過中樞神經機制誘導外周血管舒張和通過刺激迷走神經來降低血壓和心率。
5-HT1A受體激動劑如丁螺環酮和福美沙龍顯示出緩解焦慮和抑鬱的功效,並且丁螺環酮和坦度螺酮目前被批准用於世界各地的這些適應症。一些非典型抗精神病藥如阿立哌唑也是5-HT1A 受體的部分激動劑,並且有時以低劑量用作抗抑鬱藥如選擇性5-羥色胺再攝取抑製劑(SSRI)的增強劑。
通過血清素前體補充,5-羥色胺再攝取抑制,或單胺氧化酶抑制,增加5-羥色胺水平,已被證明是大多數主流抗抑鬱藥,有治療益處補充劑的主要介體。藥物包括5-羥色胺前體如L-色氨酸和5-HTP,選擇性5-羥色胺再攝取抑製劑(SSRIs),血清素-去甲腎上腺素再攝取抑製劑(SNRIs),三環抗抑鬱藥(TCAs),四環抗抑鬱藥(TeCAs)和單胺氧化酶抑製劑(MAOI)。5-HT1A 受體活化可能在血清素釋放劑(SRAs)如搖頭丸MDMA(“Ecstasy”)的作用中起重要作用。
已經證明5-HT1A 受體的激活通過抑制腦的各個區域中的谷氨酸和乙酰膽鹼的釋放而削弱記憶和學習。已知5-HT1A 激活可改善前額皮質相關的認知功能。相反,5-HT1A 體拮抗劑例如lecozotan顯示促進囓齒動物中的某些類型的學習和記憶,因此正在開發作為阿爾茨海默病的新治療藥物。
已經在科學研究中觀察到的5-HT1A 活化的其它作用包括:
減少侵略性
增加社交性
降低衝動
抑制尋求藥物的行為
促進性驅動和喚醒性
抑制陰莖勃起
減少食物攝入
延長REM睡眠潛伏期
逆轉阿片類物質誘發的呼吸抑制
內分泌學上5-HT1A 受體活化誘導各種激素的分泌,包括皮質醇,皮質酮,促腎上腺皮質激素(ACTH),催產素,催乳素,生長激素和β-內啡肽。與5-HT5 受體不同,受體不影響血管加壓素或腎素分泌。已經提出催產素釋放可能有助於在受體活化時觀察到的親社會性,抗惡化和抗焦慮性質。β-內啡肽分泌可能有助於抗抑鬱,抗焦慮和鎮痛作用。
配體
使用放射性配體碳11[11C] WAY-100,635的正電子發射斷層掃描可以對人腦中5-HT1A 受體的分佈進行成像。
部分激動劑:
阿達木黴素 安非他明 阿立哌唑 阿塞那平 蟾毒色胺 丁螺環酮 大麻二酚 氯氮平 二氫麥角胺 依托拉嗪 麥角胺 托吡酮 氟班色林 銀杏 吉哌隆 氟哌啶醇 拉莫三嗪 伊沙匹隆 鳶尾 魯拉西 麥角酸二乙酰胺(LSD) 搖頭丸(MDMA) 哌甲酯 奈法唑酮 奧氮平 吡螺酮 脫磷酸裸蓋菇素 迷幻藥 喹硫平 蘿芙素 沙立佐坦 坦度螺酮 噻洛普酮 吡唑酮 三氟甲基苯基哌嗪 烏拉地爾 維拉佐酮 育亨賓 扎洛司比 齊拉西酮等等。
完全激動劑
8-OH-DPAT Alnespirone Befiradol Eptapirone F-15,599 Lesopitron MKC-242 LY-293,284 鹽酸奧沙莫唑坦 Repinotan U-92,016-A 氟班色林等等。
拮抗劑
AV-965 BMY-7,378 氰基吲哚洛爾 Dotarizine Flopropione GR-46,611 碘代吲哚洛爾 Isamoltane Lecozotan Mefway 代謝物 甲基麥角酸 MPPF NAN-190 氧烯洛爾 Pindobind Pindolol 普萘洛爾 利培酮 Robalzotan SB-649,915 SDZ-216,525 Spiperone 螺旋藻 螺惡草 UH-301 WAY-100,135 WAY-100,635 甲苯脒等等。
5-HT2A 受體
5-HT2A 受體 該受體首先作為血清素能迷幻藥物如LSD的靶標而受到注意。後來則是許多抗精神病藥物,特別是非典型精神病的藥物而有聲譽。通過慢慢給於選擇性血清素再攝取抑製劑(SSRI)和經典抗精神病藥,可以向下調控(減少)突觸後5-HT2A 受體。自殺者與抑鬱症患者比正常患者俱有更多的5-HT2A受體。這些研究結果表明突觸後5-HT2A 密度過高參與了抑鬱症的發病機制。
分佈:5-HT2A 受體分佈於整個中樞神經系統(CNS)中。這大部分是血清素終端豐富的地區,包括大腦皮層(主要是前額葉,頂葉和體感皮層)和嗅結節區域。 特別是這種高濃度的受體在大腦皮質第五層中錐體細胞(Pyramidal cell)的頂層樹突上,可以調節認知過程,工作記憶,並通過增強谷氨酸釋放與5-HT1A ,GABAA,腺苷A1,AMPA,,mGlu5和OX2受體相互作用。在周邊中,血小板和許多心血管系統細胞中,成纖維細胞和周圍神經系統的神經元中均高度存在。
5-HT2A 受體可影響
中樞神經系統:興奮神經元,影響行為,學習,焦慮
在胃腸道和支氣管中使平滑肌收縮
使血管收縮或使血管舒張
使血小板凝集
用DOI激活5-HT2A 受體在幾種組織包括心血管和腸中產生抗炎效果。其它5-HT2A 激動劑如致幻劑(LSD)也具有針對TNF-α誘導的炎症的有效抗炎效果。
在丘腦5-HT2A 受體的活化引起催產素,催乳素,促腎上腺皮質激素,皮質酮和腎素激素濃度增加。
對記憶有角色作用
配體
迷幻劑(激動劑↑;拮抗劑↑); 抑鬱症(激動劑和拮抗劑↓); 焦慮(拮抗劑↓); 精神分裂症的陽性和陰性症狀(拮抗劑↓); 去甲腎上腺素從斑點釋放(拮抗劑↑); 膀胱收縮(激動劑↑)
激動劑
做為“經典”的致幻劑麥角二乙醯胺(LSD),脫磷酸裸蓋菇素和酶斯卡靈,必需激活5-HT2A受體,作為該受體的完全或部分激動劑,麥角靈,色胺和苯乙胺是三個主要類別,分別代表了5-HT2A激動劑。已經開發了來自這三類的非常大的衍生物家族,並對其結構 - 活性關係進行了廣泛研究。位於前額葉皮層區域內的錐體細胞的頂端樹突上,作用於5-HT2A受體的激動劑,被認為它調控致幻活動。
25I-NBOMe(完全激動劑)2C-B 5-MeO-DMT BZP 蟾毒色胺 二甲基胺(DMT) DOM 麥角新鹼 鳶尾 LSD 麥司卡林 肉荳蔻苷 PNU-22394(部分激動劑) 脫磷酸裸蓋菇素 裸蓋菇素 三氟甲基苯基哌嗪(部分激動劑或拮抗劑)
拮抗劑
非典型抗精神病藥 氯氮平 奧氮平 喹硫平 維思通 齊拉西酮 阿立哌唑 阿塞那平 阿米替林 氯米帕明 賽庚啶 Eplivanserin 托吡酮 氟哌啶醇 羥嗪 伊潘立酮 Ketanserin 甲基麥角酸 米塞林 米氮平 奈法唑酮 Pimavanserin 苯噻啶 利坦色林 曲唑酮 育亨賓
5-HT2C 受體
5-HT2C 受體是血清素的許多結合位點之一。通過血清素激活該受體抑制大腦某些區域中的多巴胺和去甲腎上腺素釋放。5-HT2C 受體顯著調節情緒,焦慮,進食和繁殖行為。5-HT2C受體調節紋狀體,前額皮質,伏隔核,海馬,下丘腦和杏仁核中的多巴胺釋放等。
研究表明,一些自殺受害者在前額葉皮層中具有異常高數量的5-HT2C 受體。有一些混合證據表明阿戈美拉汀,5-HT2C拮抗劑,是一種有效的抗抑鬱藥。 阿戈美拉汀對5-HT2C 受體的拮抗作用導致額葉皮層中多巴胺和去甲腎上腺素活性的增加。相反,許多選擇性血清素再攝取抑制劑(SSRI)通過增加突觸中血清素的水平間接刺激5-HT2C活性,儘管通常典型的選擇性血清素再攝取抑制劑(SSRIs)延遲情緒抬高通常與5-HT2C受體的減少並行。許多非典型抗精神病藥阻斷5-HT2C 受體,但是它們的臨床使用受到對各種神經遞質和受體的多種不良作用的限制。氟西汀作為直接的5-HT2C 拮抗劑,除了抑制血清素再攝取,然而該作用的臨床意義是可變動的。
HT2C 受體的過度活性可能造成某些患者群體的抑鬱和焦慮症狀。選擇性血清素再攝取抑制劑(SSRI)和血清素和去甲腎上腺素再攝取抑製劑(SNRI)藥物,例如舍曲林,帕羅西汀,文拉法辛等,引起血清素激活HT2C 受體的許多負面副作用。由SSRIs引起的一些最初的焦慮是由於在5-HT2C的過量信號傳導。 在1-2週的時間內,受體開始下調,伴隨著HT2A,HT1A和其他血清素受體的下調。這種下調與選擇性血清素再攝取抑制劑的臨床益處的發生相似。
5-HT2C 受體介導反應許多藥物對細胞外多巴胺的釋放和增加(包括咖啡因,尼古丁,苯丙胺,嗎啡,可卡因等)。
內分泌學
血清素主要通過受體亞型5-HT2A和5-HT2C的作用參與下丘腦和垂體腺激素如催乳素,促腎上腺皮質激素,加壓素和催產素的基礎和應激誘導的調控。因此,5-HT2C 受體是下丘腦-垂體-腎上腺軸(HPA軸)的重要調節劑。HPA軸是為戰鬥或逃跑反應相關的急性交感神經應激反應主控制器。長時間激活和HPA軸的擾動,有助於在許多精神病理條件下發生抑鬱症和焦慮症狀。
配體
激動劑
A-372,159 AL-38022A CP-809,101 芬氟拉明 鳶尾 氯卡色林 美舒麥角 MK-212 萘基異丙胺 諾氟氟胺 Org 12,962 ORG-37,684 Oxaflozane PNU-22394 PNU-181731 迷幻藥物 麥角酰胺(LSD等)苯乙胺(2C-B,DOI,DOM,美斯卡靈等)哌嗪(mCPP,TFMPP等)色胺(5-MeO-DMT,Bufotenin,DMT,Psilocin等)Ro60-0175 Vabicaserin WAY-629 WAY-161,503 WAY-163,909 YM-348 氯卡色林(調節食慾和食物消耗的抗肥胖剂)
部分激動劑 阿立哌唑
拮抗劑
阿戈美拉汀 CEPC 依托拉嗪 托吡酮 氟西汀 FR-260,010 Lu AA24530 甲基麥角酸 奈法唑酮 諾氟西汀 O-去甲基曲馬多 RS-102,221 SB-200,646 SB-221,284 SB-242,084 SDZ SER-082 曲馬多 曲唑酮
反激動劑
抗抑鬱藥 三環類(阿米替林,氯米帕明,丙咪嗪,去甲替林,多塞平等)四環類(米爾塔扎平,米安舍林,阿莫沙平等)抗組胺藥(Cyproheptadine,Hydroxyzine,Latrepirdine等)抗精神病藥 典型(氯丙嗪,氟奮乃靜,洛沙平,硫利達嗪等)非典型(氯氮平,奧氮平,喹硫平,利培酮,齊拉西酮等)Cinanserin Deramciclane 酮色 LY-53,857 甲麥角林 美賽 苯噻啶 利坦色林 S-32212 SB-206,553 SB-228,357 SB-243,213 SB-242,084
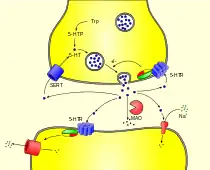
終止血清素作用
血清素作用的終止主要通過從突觸處吸收血清素。這是通過特定的單胺轉運蛋白,對在突觸前神經元對血清素、血清素轉運體(SERT),來完成這一步。各種不同藥劑可以抑制血清素再吸收,包括可卡因,右美沙芬(鎮咳藥),三環抗抑鬱藥和選擇性血清素再吸收抑製劑(SSRI)。華盛頓大學2006年進行的一項研究表明,一種新發現的單胺轉運蛋白,稱為血漿膜單胺轉運蛋白(PMAT),可能佔「血清素清除率的很大百分比」。
神经系统
中缝核神经元在大脑中5-羟色胺释放的主要来源[8]。
顯微解剖
血清素釋放到神經元之間的空間(突觸),並擴散在相對寬的間隙(>20微米),以激活位於相鄰神經元樹突,胞體和突觸前末梢的血清素受體。
當人類聞到食物時,釋放多巴胺以增加食慾。但是不同於蠕蟲,血清素並不增加人類的預期行為; 相反消耗時釋放的血清素激活多巴胺產生細胞上的HT2C受體。這停止了釋放多巴胺,從而血清素減少食慾。阻斷HT2C受體的藥物,使得身體不能夠在不再飢餓或不需要營養物時進行識別,並與體重增加相關,特別是在具有低數目受體的人羣中。海馬中HT2C受體的表現是遵循晝夜節律,正如血清素在腹內側核釋放一樣,其特徵在於在早晨時形成高峰,當時有最強的吃的動機。
的動物取得多少食物不僅僅取決於糧食可用性,而且還取決於動物與其他動物的競爭能力。這對於社會動物尤其如此,其中更強的個體可從較弱的竊取食物。因此血清素不僅參與糧食供應的可用性,也參與了社會等級的分配。
在獼猴中,領袖型獼猴(α雄性獼猴)具有比次級雄性和雌性獼猴兩倍於腦中釋放的血清素的濃度。優越狀態和腦血清中血清素濃度的高低似乎是正相關。當優勢男性從這樣的群體中除去,從屬男性開始有競爭優勢。一旦新的優越等級體系建立,新的優越個體的血清素濃度也增加到下屬男性獼猴和女性獼猴的兩倍(血清素的濃度通過腦脊液中5-羥基吲哚乙酸(5-HIAA)的水平測量)。
在人類大腦中5-HT1A受體活躍化的程度與侵略性格呈負相關,以及5-HT2A受體基因編碼突變,可能使那些具有該突變基因型的人,自殺風險加倍。腦中的血清素通常在使用後並不降解,但通過血清素轉運體細胞表面上的血清素轉運蛋白收集。研究顯示焦慮相關人格中總差異的近10%取決於描述的變化,在何處,何時和多少血清素轉運體神經元應部署。
消化道
腸道由腸嗜鉻細胞包圍,食物進管腔中細胞會釋放血清素。這使腸子圍繞食物周圍而收縮。在靜脈中的血小板吸收腸道過量的血清素。
如果食物中存在刺激物,腸嗜鉻細胞釋放更多的血清素,使腸道蠕動更快引起腹瀉,因此腸道清空了有毒物質。如果血清素在血液中釋放的速度比血小板吸收它更快,血液中游離血清素的濃度就增加。這激活化學感受器觸發區中的5-HT3 受體,引起嘔吐。腸嗜鉻細胞不僅對壞掉的食物反應,而且對輻射和癌症化療也非常敏感。阻斷5-HT3 受體的藥物在控制癌症治療產生的噁心和嘔吐方面非常有效。
骨代謝
在人類和小鼠中,血清素的濃度和信號傳導的改變顯示可以調節骨量。腦部缺乏血清素的小鼠骨質減少,腸系統缺乏血清素的小鼠骨密度高。在人類中,增加血液血清素的濃度已經顯示為低骨密度負預測物。雖然非常少,血清素也可以在骨細胞中合成。
器官發育
血清素影響器官發育。
心血管生長因子
某些疾病,例如中j腸的胃腸類癌瘤,有時釋放大量的血清素,可產生主要是右側心臟纖維化的特徵模式。這種病理學也在某些西非部落中看到,他們吃含有過量血清素的食物,如(Matoke)一種烏干達綠色香蕉。
缺乏症状
很多健康问题与大脑血清素水平低有关。造成血清素减少的原因有很多,包括压力、缺乏睡眠、营养不良和缺乏锻炼等。在降低到需要数量以下时,人们就会出现注意力集中困难等问题,会间接影响个人计划和组织能力。这种情况还经常伴随压力和厌倦感,如果血清素水平进一步下降,还会引起抑郁。
其他一些与大脑血清素水平降低有关的问题还包括易怒、焦虑、疲劳、慢性疼痛和焦躁不安等。如果不采取预防措施,这些问题会随时间推移而恶化,并最终引起强迫症、慢性疲劳综合征、关节炎、纤维肌痛和轻躁狂抑郁症等疾病。患者可能会出现不必要的侵略行为和情绪波动。血清素水平较低的人群更容易发生抑郁、冲动行为、酗酒、自杀、攻击及暴力行为,科学家甚至通过改变实验动物脑内血清素水平,使他们更具有攻击性。
代谢
血清素可以经单胺氧化酶(MAO)催化成5-羟色醛以及5-羟吲哚乙酸而随尿液排出体外。5-HT的代谢主要是通过转运体进行再摄取,一部分被降解另一部分被重新摄入囊泡。他的降解过程:经线粒体上的MAO,氧化脱氨基形成5羟吲哚乙醛,在经过醛脱氢酶的作用形成5-羟吲哚乙酸(5HILL)。5-羟吲哚乙酸可以作为抑郁症患者自杀行为的预测标记。
藥理
幾類藥物針對血清素系統,包括一些抗抑鬱藥,抗精神病藥,抗焦慮藥,止吐藥和抗偏頭痛藥物,以及迷幻藥和神入感激發劑( Empathogen)與放心藥(Entactogen)。
迷幻藥
迷幻藥脫磷酸裸蓋菇素(psilocin)/裸蓋菇素(psilocybin),二甲基胺(DMT),仙人掌毒鹼(mescaline),麥角酸二乙胺(LSD)是激動劑,主要在血清素2A/2C受體。該神入感激发剂:(Empathogen)与放心药(Entactogen), MDMA(搖頭丸)從神經元的突觸小泡釋放血清素。
以脫磷酸裸蓋菇素(psilocin)為主要成分的精神藥物商品名為賽洛新。
抗抑鬱藥
改變血清素血液濃度的藥物可用於治療抑鬱症,廣泛性焦慮症和社交恐怖症。
血清素綜合徵
血清素在血中濃度極高時可引起稱為血清素綜合徵的病症,具有毒性和潛在致死的可能。
抗精神病藥
非典型抗精神病藥物,例如氯氮平,奧氮平和利培酮(理思必妥),用於精神分裂症。利培酮是一种选择性单胺能拮抗剂,它与5-羟色胺能的5-HT2受体和多巴胺能的D2受体有很高的亲和力,也能与α1-肾上腺素能受体结合,与H1-组胺受体和α2肾上腺素能受体的亲和力较低,但不能与胆碱能受体结合。利培酮是强有力的D2拮抗剂,这是它抑制精神分裂症阳性症状的原因,但它引起的运动功能抑制及强直性昏厥都要比经典抗精神分裂症药要少,对中枢系统的5-羟色胺和多巴胺拮抗作用的平衡可以减轻发生锥体外系反应,并将其治疗作用扩展到精神分裂症的阴性症状和情感症状。
安眠藥
由於血清素對睡眠的幫助,血清素前體色氨酸和5-羥色氨酸,可幫助輕度睡眠障礙。它們的前藥其主體可以穿過血腦屏障以接納和被代謝成血清素。
鎮靜劑
丁螺環酮(Buspirone)(5-HT1A-受體的部分激動劑)也用作治療廣泛性焦慮症的精神藥物。
抑制食慾劑
血清素抑制食慾的作用是在不同的方式抑制食慾的利用。藥物如氯卡色林(Lorcaserin)和氟拉明。
偏頭痛
偏頭痛劑舒馬曲坦。
抗高血壓藥
作為抗高血壓藥,血清素拮抗劑 5-HT2A-Antagonist 酮色林(Ketanserin)和5-HT1A激動劑 5-HT1A-Agonist 烏拉地爾(Urapidil),用於治療升高的血壓。然而它們的抗高血壓作用主要不是與5-羥色胺受體的相互作用有關,而是與腎上腺素受體的相互作用有關。
抗凝劑
對5-HT2A-拮抗劑 沙格雷酯(Sarpogrelat)是一種血小板抑製劑,使用如血液抗凝劑。
止吐藥
一些血清素3拮抗劑(5-HT3 receptor antagonist),例如昂丹司瓊(ondansetron),格拉司瓊(granisetron)和托烷司瓊(tropisetron)是重要的止吐劑。 它們治療使用細胞毒性藥物的抗癌化療期間發生的噁心和嘔吐特別重要。 另一個應用是治療手術後噁心和嘔吐。
消化管機能改善薬
5-HT4-受體 西沙必利(Cisapride)為一種上胃腸道蠕動促進劑。用於胃輕癱:上消化道不適、胃-食管反流、與運動功能失調有關的假性腸梗阻、慢性便秘病人的長期治療。
其他
一些血清素激動劑藥物可在體內任何地方引起纖維化,特別是腹膜後纖維化綜合徵,以及心臟瓣膜纖維化。有三組血清素能藥物已經與這些綜合徵流行病學相關。這些是血清素能血管收縮抗偏頭痛藥物(麥角胺和甲麥角新鹼),血清素能食慾抑製劑藥物(芬氟拉明,氯芬特明和氨甲蝶呤)和某些抗帕金森病多巴胺能激動劑,它們也刺激血清素能5-HT2B受體。
甲基色胺和致幻劑
幾種植物含有血清素以及在氨基(NH2)和羥基(OH)基團處甲基化的相關色胺家族,是N-氧化物,或缺少OH基團。實例是來自柯拉豆屬(Anadenanthera genus)的植物,其用於致幻性的yopo鼻煙中。這些化合物廣泛存在於許多植物的葉中,並且可以用作動物攝取的威懾物。 血清素也發現在花褶傘屬(genus Panaeolus)的幾種蘑菇中。






比較和演化生物學
單細胞生物
各種單細胞生物體分泌血清素用於各種目的。已發現選擇性血清素再攝取抑製劑(SSRIs)對藻類有毒。胃腸道寄生蟲阿米巴分泌血清素,在一些患病者中,引起持續的分泌性腹瀉。感染溶組織阿米巴的病人,發現血清中血清素的濃度昇高,而在感染治療好後恢復正常。溶組織阿米巴也通過血清素的存在變得更有毒性。
植物



血清素存在於蘑菇,水果和蔬菜中。在核桃和山核桃屬的堅果中發現為25-400毫克/公斤(最多)。在大蕉,菠蘿,香蕉,奇異果,李子和番茄中發現5-30毫克/公斤的血清素。在廣泛的測試的蔬菜中發現了0.1-3毫克/公斤的血清素(中等水平)。
血清素是蕁麻中含有的一種有毒化合物,其在注射時會引起疼痛。它也發現在紅海扇中。
已經在巧克力中發現血清素和色氨酸。在具有85%可可的巧克力中發現最高的血清素含量(2.93微克/克),並且在70-85%可可中發現最高的色氨酸含量(13.27-13.34微克/克)。
無脊椎動物
血清素在大多數動物的神經系統中作為神經遞質的作用。例如餵養細菌時的秀麗隱桿線蟲(Caenorhabditis elegans)中,血清素作為反應陽性事件的信號而釋放,例如找到新的食物來源或在雄性動物找到一個與之交配的女性時釋放。當一個餵養良好的蠕蟲感覺細菌在其角質層會釋放多巴胺,這減慢了它;如果飢餓時也釋放血清素,這進一步減慢它。這種機制增加了動物在食物存在下所花費的時間。釋放的血清素激活了用於餵養的肌肉,而章魚胺抑制它們。血清素擴散到血清素敏感性神經元,控制動物的營養素供應的感覺。
如果龍蝦注射血清素,他們的行為像主導的個人,而注射章魚胺則引起下屬行為。淡水龍蝦那就是害怕而逃跑,並且血清素對這種行為的影響在很大程度上取決於動物的社會地位。血清素抑制下屬的逃避反應,但增強它在社會主導或個人的孤立。原因是社會經驗改變血清素受體(5-HT受體)具有對戰鬥或逃跑反應有相反的效果。5-HT1受體的作用在下屬動物中佔優勢,而5-HT2主導動物中佔優勢。
昆蟲
血清素是進化是保守的,它在昆蟲中的作用類似於人類的中樞神經系統,例如記憶,食慾,睡眠和行為。蝗蟲群由血清素介導,通過改變社會偏好和厭惡,使得凝聚成一個群居狀態。蒼蠅和蜜蜂的學習也受到血清素的影響。昆蟲血清素受體與脊椎動物有相似的序列,但藥理學上有差異。無脊椎動物藥學反應遠遠小於哺乳動物。這說明不同物種可選擇不同殺蟲劑的潛力。
黃蜂、大黃蜂和蝎子在其毒液中有血清素。如果蒼蠅餵食血清素,他們就更具侵略性。
生長繁殖
在秀麗隱桿線蟲,人工消耗血清素或增加章魚胺,典型的低食物環境:線蟲變得更活躍,並且抑制交配和產卵行為,而如果增加血清素或減少章魚胺則有相反的行為。血清素是正常雄性線蟲交配行為所必需的,其傾向離開食物尋找一個伴侶。
衰老與年齡相關的表型
已知血清素調節衰老,學習和記憶。第一個證據來自對秀麗隱桿線蟲長壽的研究。在老化的早期階段,血清素的水平增加,這改變運動行為和相關記憶。該效應通過抑制血清素受體的突變和藥物來恢復。該觀察結果與在哺乳動物和人類中血清素水平下降的觀點並不矛盾,其通常在衰老的晚期而不是在早期階段中觀察到。
缺乏與不足
基因改變的秀麗隱桿線蟲,缺乏血清素可增加生殖壽命。在休眠的幼蟲狀態可能變得肥胖,並且有時存在抑制發展現象。
參考資料
- Mazák, K.; Dóczy, V.; Kökösi, J.; Noszál, B. . Chemistry & Biodiversity. 2009, 6 (4): 578–90. PMID 19353542. doi:10.1002/cbdv.200800087.
- Erspamer, Vittorio. . Ricerca Scientifica. 1952, 22: 694–702.
- Tammisto, Tapani. . Annales medicinae experimentalis et biologiae Fenniae. 1967, 46 (3, Pt. 2): 382–4.
- King MW. . The Medical Biochemistry Page. Indiana University School of Medicine. [2009-12-01].
- Berger M, Gray JA, Roth BL. . Annu. Rev. Med. 2009, 60: 355–66. PMID 19630576. doi:10.1146/annurev.med.60.042307.110802.
- Kang K, Park S, Kim YS, Lee S, Back K. . Appl. Microbiol. Biotechnol. 2009, 83 (1): 27–34. PMID 19308403. doi:10.1007/s00253-009-1956-1.
- Francken BJ, Jurzak M, Vanhauwe JF, Luyten WH, Leysen JE. . Eur J Pharmacol. 1998, 361 (2–3): 299–309. PMID 9865521. doi:10.1016/S0014-2999(98)00744-4.
- Frazer, A.; and Hensler, J. G. . Siegel, G. J. (编). . Agranoff, Bernard W.; Fisher, Stephen K.; Albers, R. Wayne; Uhler, Michael D. Sixth. Lippincott Williams and Wilkins. 1999. ISBN 0-397-51820-X.
In 1964, Dahlstrom and Fuxe (discussed in [2]), using the Falck-Hillarp technique of histofluorescence, observed that the majority of serotonergic soma are found in cell body groups, which previously had been designated as the Raphe nuclei.
- (PDF). [2017-01-22]. (原始内容 (PDF)存档于2017-02-02).
- 提煉二甲基色胺
- List of Acacia species known to contain psychoactive alkaloids
延伸閱讀
- Gutknecht L, Jacob C, Strobel A, Kriegebaum C, Müller J, Zeng Y, Markert C, Escher A, Wendland J, Reif A, Mössner R, Gross C, Brocke B, Lesch KP. . The International Journal of Neuropsychopharmacology. June 2007, 10 (3): 309–20. PMID 17176492. doi:10.1017/S1461145706007437.
參看
- 哈尔明碱
- 選擇性5-羥色胺再攝取抑制劑(SSRI)
外部連結
| 维基共享资源中相关的多媒体资源:血清素 |
- 5-Hydroxytryptamine MS Spectrum
- Serotonin bound to proteins in the PDB
- PsychoTropicalResearch Extensive reviews on serotonergic drugs and Serotonin Syndrome.
- Molecule of the Month: Serotonin at University of Bristol
- 60-Second Psych: No Fair! My Serotonin Level Is Low, Scientific American
- Serotonin Test Interpretation on ClinLab Navigator.
