二叠纪-三叠纪灭绝事件
二疊紀-三疊紀滅絕事件(英語:,簡稱:)是一個大規模物种滅絕事件,發生於古生代二疊紀與中生代三疊紀之間,距今大約2億5140萬年[1][2]。若以消失的物種來計算,當時地球上70%的陸生脊椎動物,以及高達96%的海中生物消失[3];這次滅絕事件也造成昆蟲的唯一一次大量滅絕。計有57%的科與83%的屬消失[4][5]。在滅絕事件之後,陸地與海洋的生態圈花了數百萬年才完全恢復,比其他大型滅絕事件的恢復時間更長久[3]。此次滅絕事件是地質年代的五次大型滅絕事件中,規模最龐大的一次,因此又被正式稱為大死亡(The Great Dying)[6],或是大規模滅絕之母(Mother of all mass extinctions)[7]。

二疊紀-三疊紀滅絕事件的過程與成因仍在爭議中[8]。根據不同的研究,這次滅絕事件可分為一[1]到三[9]個階段。第一個小型高峰可能因為環境的逐漸改變,原因可能是海平面改變、海洋缺氧、盤古大陸形成引起的乾旱氣候;而後來的高峰則是迅速、劇烈的,原因可能是撞擊事件、超級火山爆發[10][11]、或是海平面驟變,引起甲烷水合物的大量釋放[12]。持续时间大约6万年[13]。
年代測定
在20世紀之前,二疊紀與三疊紀交界的地層很少被發現,因此科學家們很難準確地估算滅絕事件的年代與經歷時間,以及影響的地理範圍[14]。在1998年,科學家研究浙江省長興縣煤山附近的二疊紀/三疊紀岩層,他們採用鈾-鉛測年方法,研究鋯石中的鈾/鉛比例[2],估計二疊紀-三疊紀滅絕事件的發生年代為2億5140萬年前(誤差值為30萬年),並發現此後有隨着時間持續增高的滅絕比例[1]。在滅絕事件發生時,全球各地的碳13/碳12比例極速下降約9‰[15][16][17][18][19]。因為二疊紀/三疊紀的界線難以用放射性定年法測定,科學家們多用急遽下降的碳13/碳12比例,測定岩層中的二疊紀/三疊紀交界[20]。
在二疊紀與三疊紀交界時期,陸地與海生真菌曾有急遽的增加,原因應是當時有大量的植物與動物死亡,使得真菌得以迅速繁衍[21]。因為二疊紀/三疊紀的界線難以用放射性定年法測定,也缺乏參考用的標準化石,古動物學界一度使用真菌的繁衍高峰,測定岩層中的二疊紀/三疊紀交界。但是真菌定年法的最初提倡者,指出真菌的繁衍高峰,可能是三疊紀最早期生態系統的反覆發生現象[21]。真菌的繁衍高峰理論在最近幾年遭到許多質疑,例如:最常發現的真菌孢是Reduviasporonites,在近年被發現其實是藻類化石、[22][15]、真菌的繁衍高峰似乎不是全球性現象[23][24]、在許多地層中,真菌的繁衍高峰與二疊紀/三疊紀界線並不一致[25]。藻類被誤認為真菌胞,可能代表三疊紀時期全球各地湖泊增加,而非生物的大量死亡與腐爛[26]。在2009年的一個研究,則以生物化學方式分析Reduviasporonites的組成,例如:碳同位素、氮同位素、碳/氮比例,發現Reduviasporonites的組成接近真菌[27]。
關於這次滅絕的過程、經歷時間、以及不同生物群的滅絕模式,仍然相當模糊。一些證據顯示這次滅絕事件持續約數百萬年,而在二疊紀的最後100萬年,急遽地達到高峰[25][28]。在2000年,科學家研究浙江煤山的許多富含化石的地層,藉由統計分析指出,這次滅絕事件只有一個滅亡高峰期[1]。但是,之後近年的研究指出不同的生物群在不同的時間點滅亡;舉例而言,雖然年代難以確定,介形綱(Ostracoda)與腕足動物的大規模滅絕,先後相差了72萬年到122萬年之間[29]。在格陵蘭東部的一個化石保存良好的地層,動物群的衰退集中在6萬到1萬年之間,而植物群的衰退則持續約數十萬年之久,但也顯示滅絕事件造成全面性的影響[30]。一個較早期的研究,認為當時有兩個滅絕高峰期,相隔500萬年,這段期間的生物滅絕高於正常標準;而最後的滅絕高峰,造成當時約80%的海洋生物滅亡,其餘海生生物則多在第一次的滅絕高峰與滅絕間期消失;近年仍有研究引用這個假設[31]。若根據此一理論,第一次的滅絕高峰發生於二疊紀瓜德鲁普统末期[32]。
瓜德鲁普统末期滅絕事件對海生生物的衝擊,隨者地區與物種的不同,而有不同程度的影響。舉例而言,腕足動物與珊瑚有嚴重的大量滅亡[33][34]。所有的恐頭獸類滅亡,僅有一屬繼續存活[35];在有孔蟲門中,體型大型的紡錘蟲目(Fusulinida)也僅有費伯克蜓科(Verbeekinidae)繼續存活[36]。
滅絕模式
| 滅絕的海生生物 | 滅絕屬比例 | 備註 | ||||||
|---|---|---|---|---|---|---|---|---|
| SAR超类群 | ||||||||
| 97% | 紡錘蟲目滅絕,大部分在事件發生前滅絕 | |||||||
| 99%[37] | ||||||||
| 刺胞动物门 | ||||||||
| 96% | 古生代时期的造礁主力:四射珊瑚與床板珊瑚滅絕 | |||||||
| 触手冠动物 | ||||||||
|
苔蘚動物門 |
79% | 小窗格苔蘚蟲、變口目、隱口目滅絕 | ||||||
| 96% | 正形貝目與長身貝目滅絕 | |||||||
| 100% | 数量及多样性巅峰处于寒武纪 | |||||||
| 节肢动物门 | ||||||||
| 100% | 自志留纪開始衰退,在二叠纪-三叠纪灭绝事件發生前僅有兩屬存活 | |||||||
|
板足鱟綱(俗稱海蠍) |
100% | 可能在這次滅絕事件前不久滅絕 | ||||||
| 59% | ||||||||
| 软体动物门 | ||||||||
| 100% | ||||||||
|
雙殼綱 |
59% | |||||||
| 98% | ||||||||
| 97% | ||||||||
| 棘皮動物門 | ||||||||
| 98% | 游離海百合、圓頂海百合滅絕 | |||||||
|
海蕾綱 |
100% | 可能在這次滅絕事件前不久滅絕 | ||||||
| 100% | ||||||||
| 100% | ||||||||
| 脊索动物门 | ||||||||
|
牙形石纲 |
33%[38] | |||||||
| 100% | ||||||||
海中生物
在二疊紀-三疊紀滅絕事件中,海生無脊椎動物的滅絕程度最大。在中國南部的一些富含化石的地層,在329屬的海生無脊椎動物中,有280屬在二疊紀最後兩個沉積層(以發現牙形石化石為標準)消失[1]。
根據統計分析的結果,二疊紀末海洋生物的物種數量降低,多是因為滅亡,而非物種形成的減少[39]。受到此次滅絕事件影響最嚴重的生物群,是具有碳酸鈣外殼的海洋無脊椎動物,尤其是吸收環境中二氧化碳以製造外殼的物種[40]。
在正常時期,底棲生物的滅亡率高。這次滅絕事件使底棲生物的滅亡率,達到正常時期的數倍[41][42]。以滅絕比例來看,海洋生物遭受災難性的滅亡[43][7][1][44]。
繼續存活的海生無脊椎動物則包含:腕足動物門(牠們的數量在滅絕事件後持續了一段緩慢的衰退)、菊石亞綱的齒菊石目與海百合綱(幾乎滅亡,但後來再度繁盛、多樣化)。
存活比例最高的海洋生物,通常具有良好的循環系統、複雜的氣體交換機制、以及较少的鈣化组织。而鈣化组织最多的物種,則是滅亡比例最高的生物[45][46]。以腕足動物為例,繼續存活的物種體型較小,且是多樣化生物群落中的數量稀少成員[47]。
自從二疊紀中期羅德階之後,菊石亞綱已經歷長時間的衰退,達約3000萬年之久。瓜德鲁普统(Guadalupian)末期的有限度滅絕事件,使菊石的差異性大幅減少,環境因素可能導致這次的滅絕事件。菊石的低多樣性與低差異性持續到二疊紀末滅絕事件;然而,二疊紀末滅絕事件的範圍廣泛、過程迅速。在三疊紀,菊石開始迅速多樣化,但物種之間的差異性仍然不大[48]。
在二疊紀時期,菊石的形態空間逐漸有限。在進入三疊紀的數百萬年後,菊石再度發展出原始的形態空間,但不同演化支已出現差異[49]。
陸地無脊椎動物
在二疊紀時期,陸地的昆蟲與其他無脊椎動物非常繁盛、多樣化,其中也包含地表上曾出現過的最大型昆蟲。發生在二疊紀末的滅絕事件,是目前已知惟一的昆蟲大規模滅絕事件[4],其中有八、九個昆蟲的目消失,而至少十個目的物種減少。古網翅總目(Palaeodictyopteroidea)在二疊紀中期開始衰退,這段時間可能與植物群的變化有關係。昆蟲的衰退高峰發生於二疊紀晚期,這段時間的高峰則可能與植物群的變化(由氣候變化造成)無直接關聯[7]。
在二疊紀-三疊紀滅絕事件的前後,昆蟲的化石有極大的差異。許多古生代的昆蟲多在二疊紀-三疊紀滅絕事件後消失,只有舌鞘目、Miomoptera(內翅總目的一目)、原直翅目繼續存活到中生代。外翅總目、單尾目、古網翅總目、Protelytroptera(舌鞘目的一亞目)、原蜻蜓目在二疊紀末滅亡。在一些保存狀態良好的三疊紀晚期地層中,通常可發現與現代昆蟲族群相關的化石[4]。
植物生態系統
陸地植物的化石紀錄很少見,大多數是花粉與孢子。所有大型滅絕事件對植物造成的影響,只有到科的層級[15]。若以種層級來看,減少的物種大部分可能是埋藏過程造成的假象[15]。若以優勢物種與地理分佈來看,可發現植物生態系統有重大的變動[15]。
在二疊紀/三疊紀交界時,優勢的植物群改變,許多陸地植物突然開始衰退,例如荷達樹(裸子植物)與舌羊齒(種子蕨)[50]。在二疊紀/三疊紀交界之後,原本佔優勢的裸子植物被石松門取代[51]。
一個針對格陵蘭東部沉積層中的花粉與孢子研究,顯示在滅絕事件發生前,當地有密集的裸子植物森林。在海生無脊椎動物群衰退時,這個大型森林也開始消失,而小型草本植物開始多樣化(包含石松門的卷柏與水韭)。裸子植物後來一度興盛,然後大量滅亡。在二疊紀末滅絕事件期間與之後,植物群發生數次興衰循環。優勢植物群從大型木本植物變動到草本植物,顯示當時許多的大型植物遭到環境改變的壓力。植物群落的接續與滅絕,發生於碳13/碳12比例極速下降之後,但兩者時間並不一致[52]。裸子植物森林的復原,花了約400萬到500萬年[15]。
煤層缺口
在三疊紀早期的地層中,沒有發現煤礦床,而三疊紀中期的地層中,煤礦床薄,而且品質低[53]。關於煤層缺口的成因有許多假設。有科學家認為,當時有許多新的真菌、昆蟲、以及脊椎動物出現,這些具侵略性的動物造成植物的大量死亡。但是,這些動物也因二疊紀-三疊紀滅絕事件而大量滅亡,不太可能是煤層缺口的主因[53]。有可能所有可形成煤礦的植物,在這次滅絕事件中幾乎滅亡;而在1000萬年之後,新的植物才適應潮濕、酸性的泥炭沼澤,而出現可形成煤礦的新植物[53]。非生物性因素,則包含:減少的降雨、或增加的碎屑沉積物[15]。最後,三疊紀早期的各種沉積層很少被發現,煤層的缺口可能反映了沉積層的稀少狀況。有可能形成煤礦的三疊紀早期生態系統,目前還沒有被發現[15]。例如在澳洲東部,當時可能長時間處在寒冷的氣候,這種天氣狀況適合泥炭的形成。在可以形成泥炭的植物中,有95%在這次滅絕事件中消失[54]。但是,澳洲與南極洲的煤礦床在P-Tr事件發生前,就已經消失[15]。
陸地脊椎動物
在二疊紀末期,許多陸地脊椎動物消失,而某些演化支甚至幾乎滅亡。某些存活的族系,並未長時間繼續存在,而其他族系則開始多樣化,並長期存活着。有充足證據顯示,超過⅔的陸生兩棲動物、蜥形綱、獸孔目的科在這個滅絕事件中消失。大型的草食性動物遭受嚴重的打擊。除了前稜蜥科,幾乎所有的副爬行动物滅亡;龜鱉目具有副爬行动物的特征,曾经被認為可能是從前稜蜥科的近親演化而來,后来龜鱉目被认为属于雙孔亞綱。盤龍目早在二疊紀末期前就已滅亡。由於二疊紀的雙孔亞綱化石很少,無法確定滅絕事件對雙孔亞綱所造成的影響;雙孔亞綱包含大部分的爬行動物,例如蜥蜴、蛇、鱷魚、恐龍、以及恐龍的後代鳥類[55][56]。
滅絕模式的可能解釋
滅亡程度最嚴重的海生動物,包含:可以製造鈣質外殼(例如碳酸鈣)、代謝率低、以及呼吸系統較弱的物種,尤其是鈣質海綿類、四射珊瑚、床板珊瑚、具鈣質外殼的腕足動物、苔蘚動物門、棘皮動物門,上述動物中約有81%的屬滅亡。而無法製造鈣質外殼的近親,受到的影響較小,例如海葵。具有高代謝率、呼吸系統良好、以及缺乏鈣質外殼的動物,滅亡的幅度較小;牙形石是個例外,約有33%的屬消失[38]。
這種滅絕模式,與組織缺氧產生的效應相符合。在當時,幾乎所有的大陸棚海域都有嚴重的缺氧現象,但缺氧現象無法解釋有限度的滅絕模式;因此缺氧不是造成海生生物滅絕的唯一成因。一個針對二疊紀晚期到三疊紀早期大氣層的數學模型,指出在這段期間,大氣層中的氧氣有明顯、長時期的衰退;這個衰退不因二疊紀/三疊紀交界而增加,並在三疊紀早期達到最低點,但仍高於現今的比例。由此得知,氧氣比例的衰退與滅絕模式並不一致[38]。
另一方面,海生生物的滅絕模式也與高碳酸血症(二氧化碳過高)產生的效應相符合。當二氧化碳濃度高於正常時,將對生物造成傷害:降低呼吸色素攜帶氧到組織的能力、使體液更為酸性、妨礙動物形成碳酸鈣外殼;當濃度更高時,會使生物進入昏迷狀態。除了上述反應,二氧化碳濃度高,會使生物製造鈣質外殼更加困難。與陸地生物相比,海洋生物對於二氧化碳濃度的變化相當敏感。溶解於水中的二氧化碳,是氧的28倍。對於進行呼吸作用的動物,二氧化碳的排除,與氣體經過呼吸系統膜相關;所以海洋動物會將身體內的二氧化碳保持在低濃度,低於陸地動物。若二氧化碳濃度增加,會妨礙海洋動物合成蛋白質、降低受精機率、形成不完整的鈣質外殼[38]。
目前很少發現跨越二疊紀與三疊紀交界的陸相地層,因此難以詳細研究陸地生物的滅絕/存活比例。二疊紀與三疊紀的昆蟲有顯著的不同,但是在二疊紀晚期到三疊紀早期,有個約1500萬年的昆蟲化石斷層。目前最足以研究二疊紀/三疊紀陸地脊椎動物變化的地層,位於南非的卡魯盆地(Karoo);但目前仍沒有足夠的研究可供結論[38]。
生態系統的復原
早期的研究認為在二疊紀末滅絕事件後,地表的生物迅速的復原,但這些研究多是針對在滅絕存活的先驅生物(Pioneer organisms)的研究結果,例如水龍獸。近年的大部分研究認為,當時經歷相當長的時間,生態系統才恢復以往的高度多樣性、複雜的食物鏈、與眾多生態位。在滅絕事件後,發生數次的滅絕波動,限制棲息地的復原,因此生態系統花了長時期才完全復原。近年的研究指出,地表生態系統直到進入三疊紀中期,約是滅絕事件後400萬到600萬年,才開始復原[57]。某些研究人員更指出,直到滅絕事件後3000萬年,進入三疊紀晚期,地表生態系統才完全復原[58]。

在三疊紀早期,約是滅絕事件後400萬到600萬年,陸地的植物數量不足,造成煤層缺口,使當時的草食性動物面臨食物短缺[53]。南非卡魯盆地的河流,從曲折河改變為辮狀河,顯示該地區的植被曾長時間非常稀少[59]。
三疊紀早期的主要生態系統,無論是植物、動物,或是海洋、陸地系統,優勢生物都是少數特定、且全球分佈的物種。舉例而言,獸孔目的水龍獸,這種草食性動物的數量,佔了當時陸地草食性動物約90%;以及雙殼綱的克氏蛤(Claraia)、正海扇(Eumorphotis)、蚌形蛤(Unionites )、Promylina。正常的生態系統應由眾多物種構成,並佔據者不同的棲息地與生態位[50][60]。
在滅絕事件存活下來的某些劣勢物種,數量與棲息地大幅增加。例如:雙殼綱的海豆芽(Lingula)、出現於前寒武纪的疊層石、石松門的肋木(Pleuromeia)、種子蕨的二叉羊齒(Dicrodium)。[12][61][60][62]
海洋生態系統的改變

在二疊紀-三疊紀滅絕事件發生前,接近67%的海生動物是固著動物,附著於海床生存;在中生代期間,只有約50%的海生動物是固著動物,其餘皆是可自由行動的動物。研究顯示,事件發生後,棲息在海底表層的濾食性固著動物,例如腕足動物、海百合,物種數量減少;而構造較複雜的動物,例如蝸牛、海膽、螃蟹,物種數量增加。
在二疊紀末滅絕事件發生前,同時存在複雜與單純的海洋生態系統,比例相當;生態系統經過復原後,複雜的海洋生物群落在數量上超過單純的海洋生物群落,比例相當於3/1[63];這使海生掠食動物面臨食物競爭的壓力,促成中生代海洋生物的進一步演化。
在滅絕事件發生前,雙殼綱是非常罕見的一类。在三疊紀,雙殼綱成為數量眾多、多樣化的生物群;其中的厚殼蛤(Rudists),是形成中生代時期礁岩的主要生物。部分研究人員認為雙殼綱的興起,發生於兩次滅絕高峰的間期[64]。
在滅絕事件中,海百合綱的物種數量、多樣性大幅減少,但沒有完全滅亡[65]。在滅絕事件後,海百合綱接着發生適應輻射,除了數量、棲息地增加,也發展出柔軟的腕足、具可動性[66]。
在滅絕事件后,寒武纪演化动物群所剩无几,古生代演化动物群陷入低迷,而现代演化动物群经过三叠纪的恢复后主导着海洋生態系統[67]。
陸地脊椎動物

水龍獸屬於獸孔目二齒獸類,是種體型接近豬的草食性動物,在三疊紀最早幾個時期,佔據了90%的陸地動物生態位[12]。除此之外,還有體型較小的肉食性犬齒獸類倖存,犬齒獸類是哺乳動物的祖先。根據非洲南部卡魯盆地的化石紀錄,有少數二疊紀物種也存活過二疊紀-三疊紀滅絕事件,例如獸頭亞目的四犬齒獸、犬齒獸 (Tetracynodon)、麝喙獸(Moschorhinus)、Ictidosuchoides,但數量沒有水龍獸繁盛[68]。
在三疊紀的早期,主龍類的數量遠少於獸孔目;但在三疊紀中期,主龍類取代獸孔目的優勢生態位[12]。在三疊紀晚期,主龍類的恐龍演化出現,在侏罗纪时期才成為優勢陸地動物,此后会支配陸地生态位长达1.4亿年左右。[69]。獸孔目與主龍類的興衰,與哺乳類的演化過程有關。獸孔目與隨後出現的哺乳形類,在這個時期多半演化成小型、夜行性的食蟲動物。夜間的生活習性,至少使哺乳形類發展出較高的代謝率[70]。
兩棲動物離片椎目的某些物種,在滅絕事件中幾乎滅亡,但快速復原。在三疊紀的大部分時間,蝦蟆螈(Mastodonsaurus)與長吻迷齒螈下目(Trematosauria)是主要的水生與半水生掠食動物,以其他四足類、魚類為食[71]。
陸地脊椎動物經過相當長的時間,才從二疊紀末滅絕事件復原。一位研究人員估計陸地脊椎動物花了3000萬年,直到三疊紀晚期,才恢復之前的繁盛與多樣性。此時的陸地脊椎動物包含:兩棲動物、原始主龙形下纲、伪鳄类、恐龍、翼龍目、二齒獸類、以及哺乳形類[3]。
可能的滅絕原因
關於二疊紀-三疊紀滅絕事件的發生過程,目前已有多種假設,包含劇烈與緩慢的過程;白堊紀-第三紀滅絕事件的發生過程,也有類似的假設。劇烈過程理論的成因,包含大型或多顆隕石造成的撞擊事件、連續性火山爆發、或是海床急驟釋放出大量甲烷水合物。緩慢過程理論的成因,包含海平面改變、缺氧、以及逐漸增加的乾旱氣候[12]。
陨石撞擊事件
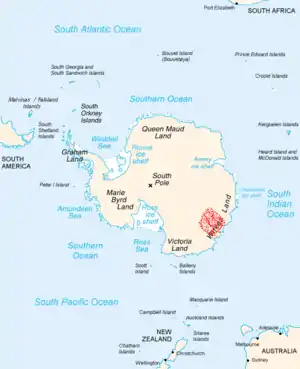
白堊紀-第三紀滅絕事件由撞擊事件造成的證據,促使科學家們推論其他滅絕事件由撞擊事件造的的可能性,尤其是二疊紀-三疊紀滅絕事件。因此,科學家們盡力尋找那個時代的大型隕石坑與撞擊證據。
目前已在部分二疊紀/三疊紀交界的地層,發現撞擊事件的證據,例如:在澳洲與南極洲發現罕見的衝擊石英[72][73]、富勒烯包覆的外太空惰性氣體[74]、南極洲發現的玻璃隕石[75],以及地層中常見鐵、鎳、矽微粒[76]。但是,上述證據的真實性多受到懷疑[77][78][79][80]。在南極洲石墨峰(Graphite Peak)發現的衝擊石英,經過光學顯微鏡與穿透式電子顯微鏡重新檢驗後,發現其中的結構並非由撞擊產生,而是形變與地殼活動(例如火山)造成的[81]。
目前已經發現數個可能與二疊紀末滅絕事件有關的隕石坑,包含:澳洲西北外海的貝德奧高地[73]、南極洲東部的威爾克斯地隕石坑[82][83]。但沒有可信服的證據,可證明這兩個地形是由撞擊產生。以威爾克斯地隕石坑為例,這個位在冰原下的凹地,年代無法確定,可能晚於二疊紀末滅絕事件才形成。其中最爭議的隕石坑,是由地質學家Michael Stanton在2002年提出的,他主張墨西哥灣是形成於二疊紀末期的一次撞擊事件,而該撞擊事件也造成了二疊紀-三疊紀滅絕事件[84]。
如果二疊紀末滅絕事件的主因是由於撞擊,很有可能隕石坑已經消失於地表。地球表面有70%是海洋,所以隕石或彗星撞擊海洋的機率,是撞擊陸地的兩倍以上。但是,地球的海洋地殼會因聚合與隱沒作用而消失於地表,所以目前無法找到距今2億年以上的海洋地殼。如果當時有非常大型的撞擊事件,撞擊會使該處地殼破裂、變薄,造成大量的熔岩[85]。
撞擊理論最受到關注的原因是,它可與其他滅絕現象產生因果連結[86],例如西伯利亞暗色岩火山爆發,可能由大型撞擊產生[87],甚至是大型撞擊的對蹠點[88][86]。即使撞擊發生於海洋,隕石坑因為隱沒帶而消失於地表,應會留下其他證據。如同白堊紀-第三紀滅絕事件,如果有撞擊事件發生,會產生大量撞擊拋出物,相同時代的許多地層會發現大量親鐵元素,例如銥。若滅絕事件由隕石撞擊引發,可以解釋在滅絕後,生物沒有快速的適應演化。
火山爆發
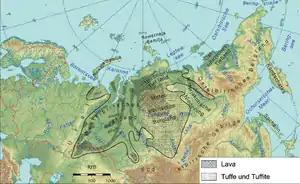
在二疊紀的最後一期,發生兩個大規模火山爆發:西伯利亞暗色岩、峨嵋山暗色岩。峨嵋山暗色岩位於現今中國四川省,規模超過30萬平方公里,形成時間是瓜德鲁普统末期,形成時的位置接近赤道[89][90]。西伯利亞暗色岩火山爆發是地質史上已知最大規模的火山爆發之一,熔岩面積超過300萬平方公里。西伯利亞暗色岩火山爆發原本被認為持續約數百萬年,但近年的研究認為西伯利亞暗色岩形成於2億5120萬年前(誤差值為30萬年),接近二疊紀末期[1][91]。
峨嵋山暗色岩與西伯利亞暗色岩火山爆發,可能製造大量灰塵與酸性微粒,遮蔽照射到地表的陽光,妨礙陸地與海洋透光帶的生物進行光合作用,進而遭成食物鏈的崩潰。大氣層中的酸性微粒,最後形成酸雨降落到地表。酸雨對陸地植物、可製造碳酸鈣硬殼的軟體動物與浮游生物造成傷害。火山爆發也釋放大量二氧化碳,形成溫室效應。大氣層中的灰塵與酸性物質降落到地表之後,過量的二氧化碳持續形成溫室效應[86]。
與其他火山相比,西伯利亞暗色岩更為危險。洪流玄武岩會產生大量的流動性熔岩,只會噴發少量的蒸氣、碎屑進入大氣層。但是,西伯利亞暗色岩火山爆發的噴出物質,似乎有20%是火山碎屑,這些火山灰與火山礫進入大氣層後,會造成短時期的氣候寒冷[92]。帶有玄武岩的熔岩侵入碳酸鹽岩、或帶有大型煤層的地區,會產生大量的二氧化碳,會在大氣層的灰塵降落到地表後,造成全球暖化[86]。
這些火山爆發事件的規模是否足以造成二疊紀末滅絕事件,仍有爭議。峨嵋山暗色岩接近赤道區,火山爆發所製造的灰塵與酸性物質,會對全世界造成影響。西伯利亞暗色岩的規模較大,但位置在北極區內,或在北極區附近。如果西伯利亞暗色岩火山爆發的持續時間在20萬年內,會使大氣層中的二氧化碳達到正常程度的兩倍。近年的氣候模型顯示,大氣層中的二氧化碳含量加倍,會使全球氣候上升1.5°C到4.5°C,這會造成嚴重的影響,但沒有到二疊紀-三疊紀滅絕事件的嚴重程度[86]。
在2005年,日本NHK與加拿大國家電影委員會(NFB)製作的《地球大進化》(;)電視節目,提出火山爆發形成的輕微全球暖化,導致甲烷水合物的氣化;由於甲烷氣體對全球暖化的影響,是二氧化碳的45倍,甲烷水合物的氣化進而導致不斷循環的全球暖化。
甲烷水合物的氣化
科學家們已在全球許多地點的二疊紀末碳酸鹽礦層中,發現當時的碳13/碳12比例有迅速減少的跡象,減少了約10‰左右[93][44]。從二疊紀末開始,碳13/碳12比例發生了一系列的上升與下降現象,直到三疊紀中期才穩定、停止;而發生於二疊紀末的第一次變動,是其中規模最大、最迅速的變動。在三疊紀中期,碳13/碳12比例穩定之後,可製造碳酸鈣外殼的動物開始復原[45]。
碳13/碳12比例的下降,可能有以下多種因素[94]:
- 火山爆發產生的氣體,其碳13/碳12比例低於正常值約5~8‰。但若要使全球的碳13/碳12比例下降約10‰,其火山爆發的規模將超越目前已知的任何地質紀錄[95]。
- 生物活動降低,使環境中的碳12更慢被攝取,更多碳12進入沉積層,而使碳13/碳12比例降低。所有的化學反應是建立在原子間的電磁力。較輕的同位素,其化學反應較快。所以生物化學過程會用到較輕的同位素。但關於古新世-始新世交替時期最大熱量(PETM页面存档备份,存于)事件的研究發現,該時期的碳13/碳12比例小幅降低約3到4‰。根據假設,即使將全部的有機碳(包含生物、土壤、海洋)進入土壤沉積層中,也不會達到古新世/始新世交替時的小幅度碳13/碳12比例下降。由此可知生物活動降低不是二疊紀末期的碳13/碳12比例降低的原因[95]。
- 埋在沉積層中的死亡生物,體內的碳13/碳12比例小於正常值約20到25‰。就理論上而言,如果海平面迅速降低,淺海地區的沉積層曝露到空氣後,開始氧化作用。但若要使全球的碳13/碳12比例下降約10‰,要有6.5到8.4兆噸有機碳經氧化後形成,而沉積層本身需要數十萬年的氧化。這似乎不太可能發生[7]。
- 間歇性海洋高氧與缺氧事件,也可能是三疊紀早期的碳13/碳12比例下降的原因[45]。全球性的海洋缺氧現象,本身也是滅絕事件的原因之一。二疊紀末期到三疊紀早期的陸地,多為熱帶地區。熱帶的大型河流會將沉積層中有有機碳帶入海洋,尤其是低緯度的鄰近海盆。生物化學過程會用到較輕的同位素,所以有機碳的碳13/碳12比例低。大量有機碳的迅速(以相對而言)釋放與沉降,可能會引發間歇性的高氧/缺氧事件[96]。元古宙晚期到寒武紀交接時期,也曾發生碳13/碳12比例的下降,可能與此相關,或是其他與海洋相關的因素[45]。

其他的理論則有:海洋大量釋放二氧化碳[97]、以及全球的碳循環系統經歷長時間的重整[94]。
但最有可能導致全球性碳13/碳12比例下降的因素,是甲烷水合物氣化產生的甲烷[7]。而利用碳循環模型模擬的結果,甲烷最有可能導致如此大幅的下降[94][97]。甲烷水合物是固態形式的水於晶格(水合物)中包含大量的甲烷。甲烷是由甲烷菌製造,碳13/碳12比例低於正常值約60‰。甲烷在特定的壓力與溫度下,會形成包合物,例如永凍層的近表層,並在大陸棚、更深的海床等地區大量形成。甲烷水合物通常出現在海平面300公尺以下的沉積層。最深可在水深2000公尺處發現,但大部分在水深1100公尺以上[98]。
西伯利亞暗色岩火山爆發產生的熔岩面積,是以往認定的兩倍以上,新發現的熔岩地區,在二疊紀末時幾乎是淺海。極有可能這些淺海地區蘊含甲烷水合物,而火山爆發產生的熔岩流入海床後,促使甲烷水合物的汽化[99]。
由於甲烷本身是種非常強的溫室氣體,大量的甲烷被視為造成全球暖化的主要原因。證據顯示該時期的全球氣溫上升,赤道區上升約6°C,高緯度地區上升更多。例如:氧18/氧16比例的下降[100]、舌羊齒植物群(舌羊齒與生存於相同地區的植物)消失,由生存於低緯度的植物群取代[101][12]。
碳13/碳12比例的變動,被認為與大量釋放的甲烷有關,但兩者在三疊紀早期的變動模式,並不吻合。要造成如此大的氣候變遷,所需要的甲烷量,是引發古新世-始新世交替時期最大熱量的五倍[45]。但三疊紀早期曾出現大量碳13,使碳13/碳12比例迅速增加,而後下降[45]。
海平面改變
當原本沉浸的海床露出海平面時,會造成海退。海平面的下降會使淺海的生存區域減少,破壞當地的生態系統。淺海的可棲息地,富含食物鍊下層的生物,這些生物的減少,使賴其以維生的生物競爭食物更激烈[102]。海退與滅絕事件之間似乎有部份關聯,但另有證據認為兩者間沒有關係,而海退會形成新的棲息地。[12]海平面的變化,同時也導致海底沉積物的變化,並影響海水溫度與鹽度,進一步造成海生生物的多樣性衰退[103]。
海洋缺氧
有證據顯示,二疊紀末期的海洋發生了缺氧事件。在格陵蘭東部的一個二疊紀末期海相沉積層,指出當時有明顯、快速的海洋缺氧現象[104]。而數個二疊紀末沉積層的鈾/釷比例,也指出在這次滅絕事件發生時,海洋有嚴重的缺氧現象[105]。
缺氧事件可能導致海洋生物的大量死亡,只有棲息於海底泥層、可以進行無氧呼吸的細菌不受影響。另有證據顯示,這次海洋缺氧事件,造成海床大量釋放硫化氫。
海洋缺氧事件的原因,可能是長時間的全球暖化,降低赤道區與極區之間的溫度梯度,進而造成溫鹽環流系統的緩慢,甚至停止。溫鹽環流系統的緩慢或停止,可能使得海洋中的含氧量減少[105]。
但是,某些研究人員架構出二疊紀末期的海洋溫鹽環流系統,認為當時的溫鹽環流系統無法解釋深海區域的缺氧現象[106]。
硫化氫
二疊紀末期發生的海洋缺氧事件,可能使硫酸鹽還原菌成為海洋生態系統中的優勢物種,包含脫硫桿菌目、脫硫弧菌目、互營桿菌目、熱脫硫桿菌門,這些生物會製造大量的硫化氫,過量的硫化氫會對陸地、海洋中的動植物造成毒害,並破壞臭氧層,使生物暴露在紫外線下[107]。在二疊紀末到三疊紀早期發現許多綠菌,它們進行不產氧光合作用,釋放出硫化氫。綠菌的興盛時期,與二疊紀末滅絕事件和事後的長期復原,時期相符[108]。大氣層中的二氧化碳增加,植物卻大規模滅亡,硫化氫理論可以解釋植物的大規模滅亡。二疊紀末地層中的孢子化石,多數帶有不正常特徵,可能是由硫化氫破壞臭氧層,大量的紫外線進入地表造成。
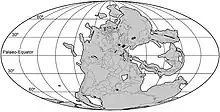
盤古大陸的形成
在二疊紀中期(約是烏拉爾世的空谷階),幾乎地表的所有大陸聚合成盤古大陸,盤古大陸由泛大洋環繞着,而東亞部份直到二疊紀末期才與盤古大陸聚合[109]。盤古大陸的完全形成,使得全球大部分的淺水區域消失,而淺水區域是海洋中最多生物棲息的部份。原本隔離的大陸架連接之後,使彼此獨立的生態系統開始互相競爭。盤古大陸的形成,造成了單一的海洋循環系統,以及單一的大氣氣候系統,在盤古大陸的海岸形成季風氣候,而廣大的內陸則形成乾旱的氣候。
在盤古大陸完全形成後,海洋生物數量開始減少,滅亡比例接近其他大型滅絕事件。盤古大陸的形成,似乎對陸地生物沒有造成嚴重的變化,二疊紀晚期的獸孔目反而因此擴大生存領域、更為多樣性。所以盤古大陸的完全形成并不是導致二疊紀末滅絕事件的直接原因。
参考文献
- (英文) Jin YG, Wang Y, Wang W, Shang QH, Cao CQ, Erwin DH. . Science. 2000, 289 (5478): 432–436. PMID 10903200. doi:10.1126/science.289.5478.432.
- (英文)Bowring SA, Erwin DH, Jin YG, Martin MW, Davidek K, Wang W. . Science. 1998, 280 (1039): 1039–1045. doi:10.1126/science.280.5366.1039.
- (英文)Benton M J. . Thames & Hudson. 2005. ISBN 978-0500285732.
- (英文)Labandeira CC, Sepkoski JJ. . Science. 1993, 261 (5119): 310–5. PMID 11536548.
- (英文)Sole, R. V., and Newman, M., 2002. "Extinctions and Biodiversity in the Fossil Record - Volume Two, The earth system: biological and ecological dimensions of global environment change" pp. 297-391, Encyclopedia of Global Enviromental Change John Wilely & Sons.
- (英文)Barry, Patrick L. . Science@NASA. Science and Technology Directorate, Marshall Space Flight Center, NASA. January 28, 2002 [March 26, 2009]. (原始内容存档于2012年2月16日).
- (英文)Erwin DH. . Columbia University Press. 1993. ISBN 0231074670.
- (英文)Yin H, Zhang K, Tong J, Yang Z, Wu S. . Episodes: 102–114.
- (英文)Yin HF, Sweets WC, Yang ZY, Dickins JM,. . Cambridge Univ. Pres, Cambridge, 1992.
- . [2020-12-15]. (原始内容存档于2020-07-29).
- . [2011-01-25]. (原始内容存档于2011-01-31).的存檔,存档日期2011-01-31.
- (英文)Tanner LH, Lucas SG & Chapman MG. (PDF). Earth-Science Reviews. 2004, 65 (1-2): 103–139 [2007-10-22]. doi:10.1016/S0012-8252(03)00082-5. (原始内容 (PDF)存档于2007-10-25).
- Burgess, S. D.; Bowring, S.; Shen, S.-z. . Proceedings of the National Academy of Sciences. 2014-02-10, 111 (9): 3316–3321. doi:10.1073/pnas.1317692111.
- (英文)Erwin, D.H. . New York isbn=0231074670: Columbia University Press. 1993.
- (英文)McElwain, J.C.; Punyasena, S.W. . Trends in Ecology & Evolution. 2007, 22 (10): 548–557. doi:10.1016/j.tree.2007.09.003.
- (英文)Magaritz, M. . Geology. 1989, 17: 337–340.
- (英文)Krull, S.J., and Retallack, J.R. . GSA Bulletin. 2000, 112 (9): 1459–1472. ISSN 0016-7606. doi:10.1130/0016-7606(2000)112<1459:CDPFPA>2.0.CO;2.
- (英文)Dolenec, T., Lojen, S., Ramovs, A. . Chemical Geology. 2001, 175 (1): 175–190. doi:10.1016/S0009-2541(00)00368-5.
- (英文)Musashi, M., Isozaki, Y., Koike, T. and Kreulen, R. . Earth Planet. Sci. Lett. 2001, 193: 9–20. doi:10.1016/S0012-821X(01)00398-3.
- (英文)Dolenec, T., Lojen, S., and Ramovs, A. . Chemical Geology. 2001, 175: 175–190. doi:10.1016/S0009-2541(00)00368-5.
- (英文)H Visscher, H Brinkhuis, D L Dilcher, W C Elsik, Y Eshet, C V Looy, M R Rampino, and A Traverse. . Proceedings of the National Academy of Sciences. 1996, 93 (5): 2155–2158 [2007-09-20].
- (英文)Foster, C.B.; Stephenson, M.H.; Marshall, C.; Logan, G.A.; Greenwood, P.F. . Palynology. 2002, 26 (1): 35–58 [2008-08-16]. doi:10.2113/0260035. (原始内容存档于2011-07-26).
- (英文)López-Gómez, J. and Taylor, E.L. . Palaeogeography, Palaeoclimatology, Palaeoecology. 2005, 229 (1-2): 1–2 [2008-08-16]. doi:10.1016/j.palaeo.2005.06.028. (原始内容存档于2010-03-06).
- (英文)Looy, C.V.; Twitchett, R.J.; Dilcher, D.L.; Van Konijnenburg-van Cittert, J.H.A.; Visscher, H. . Proceedings of the National Academy of Sciences. 2005, 162 (4): 653–659. PMID 11427710. doi:10.1073/pnas.131218098.
See image 2
- (英文)Ward PD, Botha J, Buick R, De Kock MO, Erwin DH, Garrison GH, Kirschvink JL & Smith R. . Science. 2005, 307 (5710): 709–714. doi:10.1126/science.1107068.
- (英文)Retallack, G.J.; Smith, R.M.H.; Ward, P.D. . Bulletin of the Geological Society of America. 2003, 115 (9): 1133–1152 [2008-08-16]. doi:10.1130/B25215.1. (原始内容存档于2008-05-05).
- Sephton, Mark A.; Visscher, Henk; Looy, Cindy V.; Verchovsky, Alexander B.; Watson, Jonathon S. . Geology. 2009, 37 (10): 875–878 [2010-03-07]. doi:10.1130/G30096A.1. (原始内容存档于2012-04-28).
- (英文)Rampino MR, Prokoph A & Adler A. . Geology. 2000, 28 (7): 643–646. Bibcode:2000Geo....28..643R. ISSN 0091-7613. doi:10.1130/0091-7613(2000)28<643:TOTEEH>2.0.CO;2.
- (英文)Wang, S.C.; Everson, P.J. . Paleobiology. 2007, 33 (2): 324–336. doi:10.1666/06056.1.
- (英文)Twitchett RJ Looy CV Morante R Visscher H & Wignall PB. . Geology. 2001, 29 (4): 351–354. doi:10.1130/0091-7613(2001)029<0351:RASCOM>2.0.CO;2.
- Retallack, G.J.; Metzger, C.A.; Greaver, T.; Jahren, A.H.; Smith, R.M.H.; Sheldon, N.D. . Bulletin of the Geological Society of America. 2006, 118 (11-12): 1398–1411. doi:10.1130/B26011.1.
- (英文)Stanley SM & Yang X. . Science. 1994, 266 (5189): 1340–1344. PMID 17772839. doi:10.1126/science.266.5189.1340.
- (英文)Shen, S., and Shi, G.R. . Paleobiology. 2002, 28: 449–463. doi:10.1666/0094-8373(2002)028<0449:PEPOPB>2.0.CO;2.
- (英文)Wang, X-D, and Sugiyama, T. . Lethaia. December 2000, 33 (4): 285–294. doi:10.1080/002411600750053853.
- Retallack, G.J., Metzger, C.A., Jahren, A.H., Greaver, T., Smith, R.M.H., and Sheldon, N.D. . GSA Bulletin. November/December 2006, 118 (11/12): 1398–1411. doi:10.1130/B26011.1.
- (英文)Ota, A, and Isozaki, Y. . Journal of Asian Earth Sciences. March 2006, 26 (3-4): 353–368.
- (英文) Racki G. . Palaeogeography Palaeoclimatology Palaeoecology. 1999, 154 (1–2): 107–132. doi:10.1016/S0031-0182(99)00089-9.
- (英文)Knoll, A.H., Bambach, R.K., Payne, J.L., Pruss, S., and Fischer, W.W. . Earth and Planetary Science Letters. 2007, 256: 295–313 [2008-07-04]. doi:10.1016/j.epsl.2007.02.018. (原始内容存档于2008-04-10). Full contents may be available online at (PDF). [2008-07-04]. (原始内容 (PDF)存档于2020-10-04).
- Bambach, R.K.; Knoll, A.H.; Wang, S.C., , Paleobiology, December 2004, 30 (4): 522–542 [2008-08-16], (原始内容存档于2020-05-29)
- Knoll, A.H., (PDF), (2004) [2010-03-07], (原始内容 (PDF)存档于2010-06-20)
- (英文)Stanley, S.M. . Paleobiology. 2008, 34 (1): 1–21 [2008-05-13]. doi:10.1666/07026.1. (原始内容存档于2011-07-26).
- (英文)Stanley, S.M. . Paleobiology. 2007, 33 (sp6): 1–55 [2008-08-16]. doi:10.1666/06020.1. (原始内容存档于2011-07-26).
- (英文)McKinney, M.L. . Nature. 1987, 325 (6100): 143–145. doi:10.1038/325143a0.
- (英文)Twitchett RJ, Looy CV, Morante R, Visscher H, Wignall PB. . Geology. 2001, 29 (4): 351–354. doi:10.1130/0091-7613(2001)029%3C0351:RASCOM%3E2.0.CO;2.
- (英文)Payne, J.L.; Lehrmann, D.J.; Wei, J.; Orchard, M.J.; Schrag, D.P.; Knoll, A.H. . Science. 2004, 305 (5683): 506 [2008-09-22]. PMID 15273391. doi:10.1126/science.1097023. (原始内容存档于2010-04-04).
- (英文)Knoll, A.H.; Bambach, R.K.; Canfield, D.E.; Grotzinger, J.P. . Science (Washington). 1996, 273 (5274): 452–456. PMID 8662528. doi:10.1126/science.273.5274.452.
- (英文)Leighton, L.R.; Schneider, C.L. . Paleobiology. 2008, 34 (1): 65–79. doi:10.1666/06082.1.
- (英文)Villier, L. . Science. 2004, 306: 264. doi:10.1126/science.1102127.
- (英文)Saunders, W. B. . Paleobiology. 2008, 34: 128. doi:10.1666/07053.1.
- (英文)Retallack, GJ. . Science. 1995, 267 (5194): 77–80. doi:10.1126/science.267.5194.77.
- (英文)Looy, CV Brugman WA Dilcher DL & Visscher H. . Proceedings National Academy of Sciences. 1999, 96: 13857–13862. PMID 10570163.
- (英文)Looy, CV; Twitchett RJ, Dilcher DL, &Van Konijnenburg-Van Cittert JHA and Henk Visscher. . Proceedings of the National Academy of Sciences. July 3, 2001, 14 (98): 7879–7883. doi:10.1073/pnas.131218098.
- (英文)Retallack GJ Veevers JJ & Morante R. . GSA Bulletin. 1996, 108 (2): 195–207 [2007-09-29]. (原始内容存档于2010-05-29).
- (英文)Michaelsen P. . Palaeogeography, Palaeoclimatology, Palaeoecology. 2002, 179 (3–4): 173–188. doi:10.1016/S0031-0182(01)00413-8.
- (英文)Maxwell, W. D. . Palaeontology. 1992, 35: 571–583.
- Erwin DH. . Annual Review of Ecology and Systematics. 1990, 21: 69–91. doi:10.1146/annurev.es.21.110190.000441.
- (英文)Lehrmann, D.J., Ramezan, J., Bowring, S.A.; 等. . Geology. December 2006, 34 (12): 1053–1056 [2008-09-24]. doi:10.1130/G22827A.1. (原始内容存档于2011-11-06).
- (英文)Sahney, S. and Benton, M.J. (PDF). Proceedings of the Royal Society: Biological. 2008, 275: 759 [2008-09-24]. doi:10.1098/rspb.2007.1370. (原始内容存档 (PDF)于2011-02-22).
- Ward PD, Montgomery DR, & Smith R. . Science. 2000, 289 (5485): 1740–1743. doi:10.1126/science.289.5485.1740.
- (英文)Hallam A & Wignall PB. . Oxford University Press. 1997. ISBN 978-0198549161.
- (英文)Zi-qiang W. . Review of Palaeobotany and Palynology. 1996, 91 (1–4): 121–142. doi:10.1016/0034-6667(95)00069-0.
- Rodland, DL & Bottjer, DJ. . Palaios. 2001, 16 (1): 95–101. doi:10.1669/0883-1351(2001)0163%C0095:BRFTEP3%E2.0.CO;2.
- (英文)Wagner PJ, Kosnik MA, & Lidgard S. . Science. 2006, 314 (5803): 1289–1292. doi:10.1126/science.1133795.
- (英文)Clapham, M.E., Bottjer, D.J. and Shen, S. . Geological Society of America Abstracts with Programs. 2006, 38 (7): 117 [2008-03-28]. (原始内容存档于2015-12-08).
- (英文)Foote, M. (PDF). Paleobiology. 1999, 25 (sp1): 1–116 [2008-05-12]. doi:10.1666/0094-8373(1999)25[1:MDITER]2.0.CO;2.
- (英文)Baumiller, T. K. . Annual Review of Earth and Planetary Sciences (PDF). 2008, 36: 221–249. doi:10.1146/annurev.earth.36.031207.124116.
- Servais, Thomas; Harper, David A.T.; Munnecke, Axel; Owen, Alan W.; Sheehan, Peter M. . GSA Today. 2009, 19 (4): 4 [2020-02-22]. ISSN 1052-5173. doi:10.1130/GSATG37A.1. (原始内容存档于2020-07-23) (英语).
- Botha, J., and Smith, R.M.H. . Lethaia. 2007, 40: 125–137 [2008-07-02]. doi:10.1111/j.1502-3931.2007.00011.x. (原始内容存档于2012-12-16). Full version online at (PDF). [2008-07-02]. (原始内容 (PDF)存档于2008-09-10).
- (英文)Benton, M.J. . Blackwell Publishers. 2004: xii–452. ISBN 0-632-05614-2.
- (英文)Ruben, J.A., and Jones, T.D. . American Zoologist. 2000, 40 (4): 585–596 [2008-08-16]. doi:10.1093/icb/40.4.585. (原始内容存档于2009-08-29).
- (英文)Yates AM & Warren AA. . Zoological Journal of the Linnean Society. 2000, 128 (1): 77–121 [2008-01-18]. (原始内容存档于2007-10-01).
- (英文)Retallack GJ, Seyedolali A, Krull ES, Holser WT, Ambers CP, Kyte FT. . Geology. 1998, 26 (11): 979–982 [2008-09-24]. (原始内容存档于2011-09-28).
- (英文)Becker L, Poreda RJ, Basu AR, Pope KO, Harrison TM, Nicholson C, Iasky R. . Science. 2004, 304 (5676): 1469–1476 [2008-09-24]. doi:10.1126/science.1093925. (原始内容存档于2010-09-19).
- (英文)Becker L, Poreda RJ, Hunt AG, Bunch TE, Rampino M. . Science. 2001, 291 (5508): 1530–1533. doi:10.1126/science.1057243.
- (英文)Basu AR, Petaev MI, Poreda RJ, Jacobsen SB, Becker L. . Science. 2003, 302 (5649): 1388–1392. doi:10.1126/science.1090852.
- (英文)Kaiho K, Kajiwara Y, Nakano T, Miura Y, Kawahata H, Tazaki K, Ueshima M, Chen Z, Shi GR. . Geology. 2001, 29 (9): 815–818 [2007-10-22]. (原始内容存档于2011-09-28).
- (英文)Farley KA, Mukhopadhyay S, Isozaki Y, Becker L, Poreda RJ. . Science. 2001, 293 (5539): 2343. doi:10.1126/science.293.5539.2343a.
- (英文)Koeberl C, Gilmour I, Reimold WU, Philippe Claeys P, Ivanov B. . Geology. 2002, 30 (9): 855–856. doi:10.1130/0091-7613(2002)030%3C0855:EPCBBI%3E2.0.CO;2.
- (英文)Isbell JL, Askin RA, Retallack GR. . Geology. 1999, 27 (9): 859–860. doi:10.1130/0091-7613(1999)027%3C0859:SFEOIA%3E2.3.CO;2.
- (英文)Koeberl K, Farley KA, Peucker-Ehrenbrink B, Sephton MA. . Geology. 2004, 32 (12): 1053–1056. doi:10.1130/G20907.1.
- (英文)Langenhorst F, Kyte FT & Retallack GJ. (PDF). . 2005 [2007-07-13]. (原始内容存档 (PDF)于2020-11-15).
- (英文)von Frese RR, Potts L, Gaya-Pique L, Golynsky AV, Hernandez O, Kim J, Kim H & Hwang J. . Eos Trans. AGU, Jt. Assem. Suppl. 2006, 87 (36): Abstract T41A–08 [2007-10-22]. (原始内容存档于2007-09-30).
- (英文)Von Frese, R.R.B.; L. V. Potts, S. B. Wells, T. E. Leftwich, H. R. Kim, J. W. Kim, A. V. Golynsky, O. Hernandez, and L. R. Gaya-Piqué. . Geochem. Geophys. Geosyst. 2009, 10: Q02014. doi:10.1029/2008GC002149.
- (英文)Is the Gulf's Origin Heaven Sent? by Michael S. Stanton Explorer magazine article "Is the Gulf's Origin Heaven Sent?" 页面存档备份,存于
- (英文)Jones AP, Price GD, Price NJ, DeCarli PS, Clegg RA. . Earth and Planetary Science Letters. 2002, 202 (3): 551–561. doi:10.1016/S0012-821X(02)00824-5.
- (英文)White RV. (PDF). Phil. Trans. Royal Society of London. 2002, 360: 2963–2985 [2008-01-12]. doi:10.1098/rsta.2002.1097. (原始内容存档 (PDF)于2020-11-11).
- (英文)AHager, Bradford H. . . 2001 [2008-09-24]. (原始内容存档于2008-04-22).
- (英文)Hagstrum, Jonathan T. . . 2001 [2008-09-24]. (原始内容存档于2008-04-22).
- (英文)Zhou, M-F., Malpas, J, Song, X-Y, Robinson, PT, Sun, M, Kennedy, AK, Lesher, CM & Keays, RR. . Earth and Planetary Science Letters. 2002, 196 (3–4): 113–122. doi:10.1016/S0012-821X(01)00608-2.
- (英文)Wignall, Paul B.; et al.. . Science. 2009, 324 (5931): 1179–1182. doi:10.1126/science.1171956.
- (英文)Mundil, R., Ludwig, K.R., Metcalfe, I. & Renne, P.R. . Science. 2004, 305 (5691): 1760–1763. doi:10.1126/science.1101012.
- (英文)"Permian–Triassic Extinction - Volcanism" 页面存档备份,存于
- (英文)Palfy J, Demeny A, Haas J, Htenyi M, Orchard MJ, & Veto I. . Geology. 2001, 29 (11): 1047–1050. doi:10.1130/0091-7613(2001)029%3C1047:CIAAOG%3E2.0.CO;2.
- (英文)Berner, R.A. . Proceedings of the National Academy of Sciences. 2002, 99: 32095199. PMID 11917102. doi:10.1073/pnas.032095199.
- (英文)Dickens GR, O'Neil JR, Rea DK & Owen RM. . Paleoceanography. 1995, 10 (6): 965–71. doi:10.1029/95PA02087.
- (英文)Schrag, D.P., Berner, R.A., Hoffman, P.F., and Halverson, G.P. . Geochemistry Geophysics Geosystems. 2002, 3 (6 doi=10.1029/2001GC000219): 1036 [2008-09-25]. (原始内容存档于2008-04-21). Preliminary abstract at Schrag, D.P. . Geological Society of America. June 2001 [2008-09-25]. (原始内容存档于2018-04-25).
- (英文)Benton, M.J.; Twitchett, R.J. . Trends in Ecology & Evolution. 2003, 18 (7): 358–365. doi:10.1016/S0169-5347(03)00093-4.
- (英文)Dickens GR. . Organic Geochemistry. 2001, 32 (10): 1179–1193. doi:10.1016/S0146-6380(01)00086-9.
- (英文)Reichow MK, Saunders AD, White RV, Pringle MS, Al'Muhkhamedov AI, Medvedev AI & Kirda NP. . Science. 2002, 296 (5574): 1846–1849. doi:10.1126/science.1071671.
- (英文)Holser WT, Schoenlaub H-P, Attrep Jr M, Boeckelmann K, Klein P, Magaritz M, Orth CJ, Fenninger A, Jenny C, Kralik M, Mauritsch H, Pak E, Schramm J-F, Stattegger K & Schmoeller R. . Nature. 1989, 337 (6202): 39–44. doi:10.1038/337039a0.
- (英文)Dobruskina IA. . Palaeogeography, Palaeoclimatology, Palaeoecology. 1987, 58 (1-2): 75–86. doi:10.1016/0031-0182(87)90007-1.
- (英文)Newell ND. (PDF). American Museum novitates. 1971, 2465: 1–37 [2007-11-03]. (原始内容存档 (PDF)于2008-12-17).
- (英文)McRoberts, C.A., Furrer, H., Jones, D.S. . Palaeogeography Palaeoclimatology Palaeoecology. 1997, 136 (1-4): 79– 95. doi:10.1016/S0031-0182(97)00074-6.
- (英文)Wignall PB & Twitchett RJ. . Journal of the Geological Society. 2002, 159 (6): 691–703. doi:10.1144/0016-764900-120.
- (英文)Monastersky, R., , Science News, May 25, 1996 [2008-09-22], (原始内容存档于2008-10-19)
- Zhang R, Follows, MJ, Grotzinger, JP, & Marshall J. . Paleoceanography. 2001, 16 (3): 317–329 [2008-09-25]. doi:10.1029/2000PA000522. (原始内容存档于2008-04-21).
- (英文)Kump LR, Pavlov A, & Arthur MA. . Geology. 2005, 33 (5): 397–400. doi:10.1130/G21295.1.
- (英文)Grice K, Cao C, Love GD, Bottcher ME, Twitchett RJ, Grosjean E, Summons RE, Turgeon SC, Dunning W & Yugan J. . Science. 2005, 307 (5710): 706–709. doi:10.1126/science.110432.
- (英文)The Permian - Palaeos 页面存档备份,存于
延伸阅读
- (英文)Over, Jess (editor), Understanding Late Devonian and Permian–Triassic Biotic and Climatic Events, (Volume 20 in series Developments in Palaeontology and Stratigraphy (2006). The state of the inquiry into the extinction events.
- (英文)Sweet, Walter C. (editor), Permo–Triassic Events in the Eastern Tethys : Stratigraphy Classification and Relations with the Western Tethys (in series World and Regional Geology)
外部連結
- 專門性:
- 二疊紀-三疊紀滅絕事件:化石證據、滅亡物種、滅絕原因與分析
- 二疊紀-三疊紀滅絕事件:海洋、陸地動物與植物的滅絕,碳、氧、鍶同位素的變化,年代測定,時間與過程模式
- 與二疊紀末滅絕事件有關的南極洲冰層證據
- 生物史上的五次滅絕事件页面存档备份,存于
- 西伯利亞暗色岩的年代、規模、成因、與影響页面存档备份,存于
- 新聞性:
- 科學網:2.5億年前兩大物種滅絕之謎獲突破性進展
- BBC:二疊紀末滅絕事件的複雜原因页面存档备份,存于
- BBC2:在2.5億年前,地球上的生物幾乎滅亡Archived 2012-08-08 at WebCite
- SpaceRef:南極洲冰層下的隕石坑(影像由俄亥俄州立大學提供)
- Science Daily:全球暖化使大氣層充滿硫化氫,並導致二疊紀末滅絕事件页面存档备份,存于
- Science Daily:南極洲的冰層下發現疑似隕石坑的大坑洞页面存档备份,存于
- SPACE.com:南極洲發現地表最大的隕石坑,可能是地質年代中最嚴重的生物滅絕有關]
- Space.com:地層中的二疊紀末生物滅絕證據页面存档备份,存于
- NAI:二疊紀滅絕事件是否由撞擊事件導致?